
Population Differentiation in Acer platanoides L. at the Regional Scale—Laying the Basis for Effective Conservation of Its Genetic Resources in Austria

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Populations Sampled
2.2. Marker Development and Genotyping
2.3. Data Analysis
3. Results
3.1. Novel Microsatellite Markers
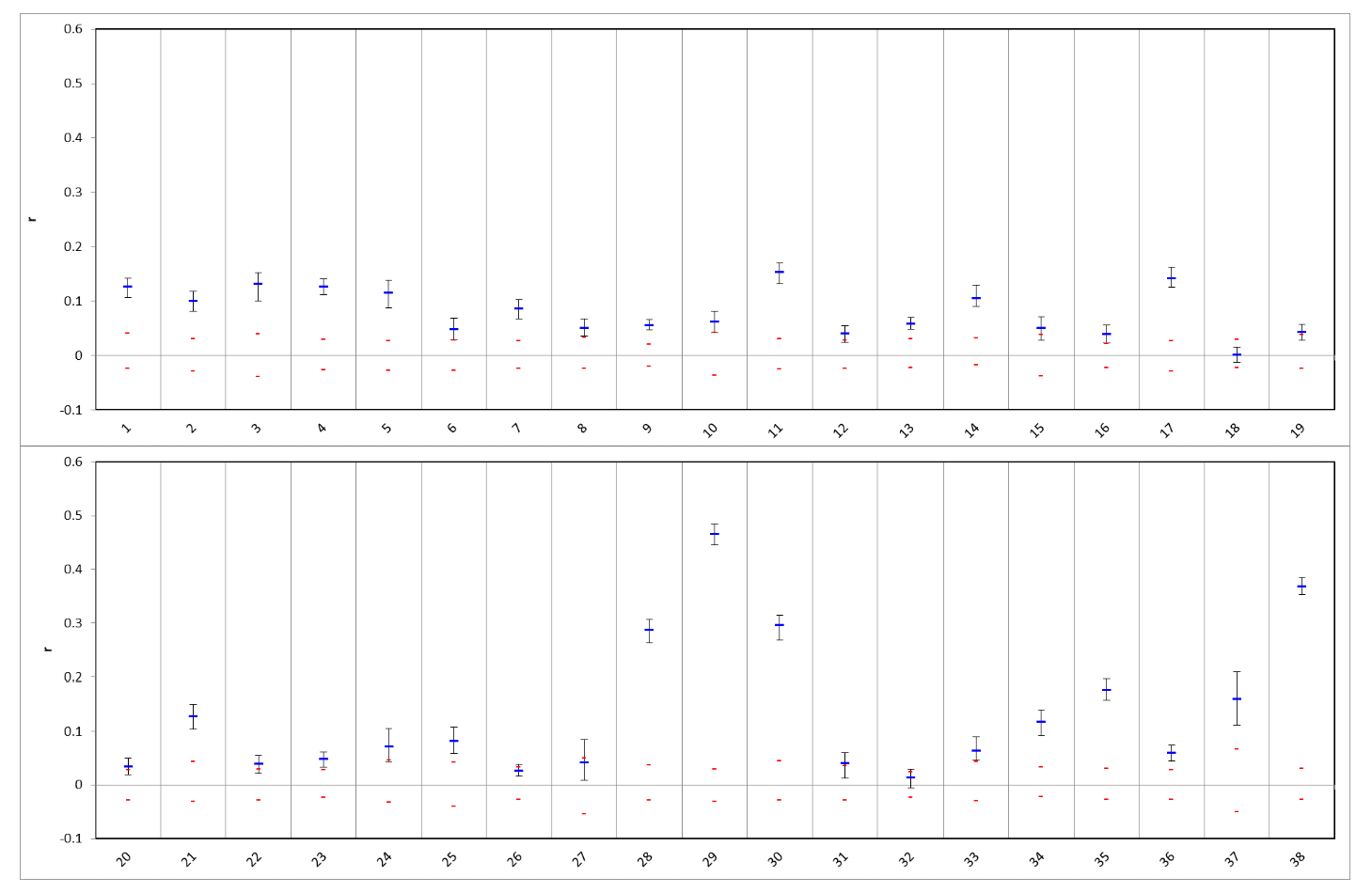
3.2. Genetic Diversity
3.3. Population Structure
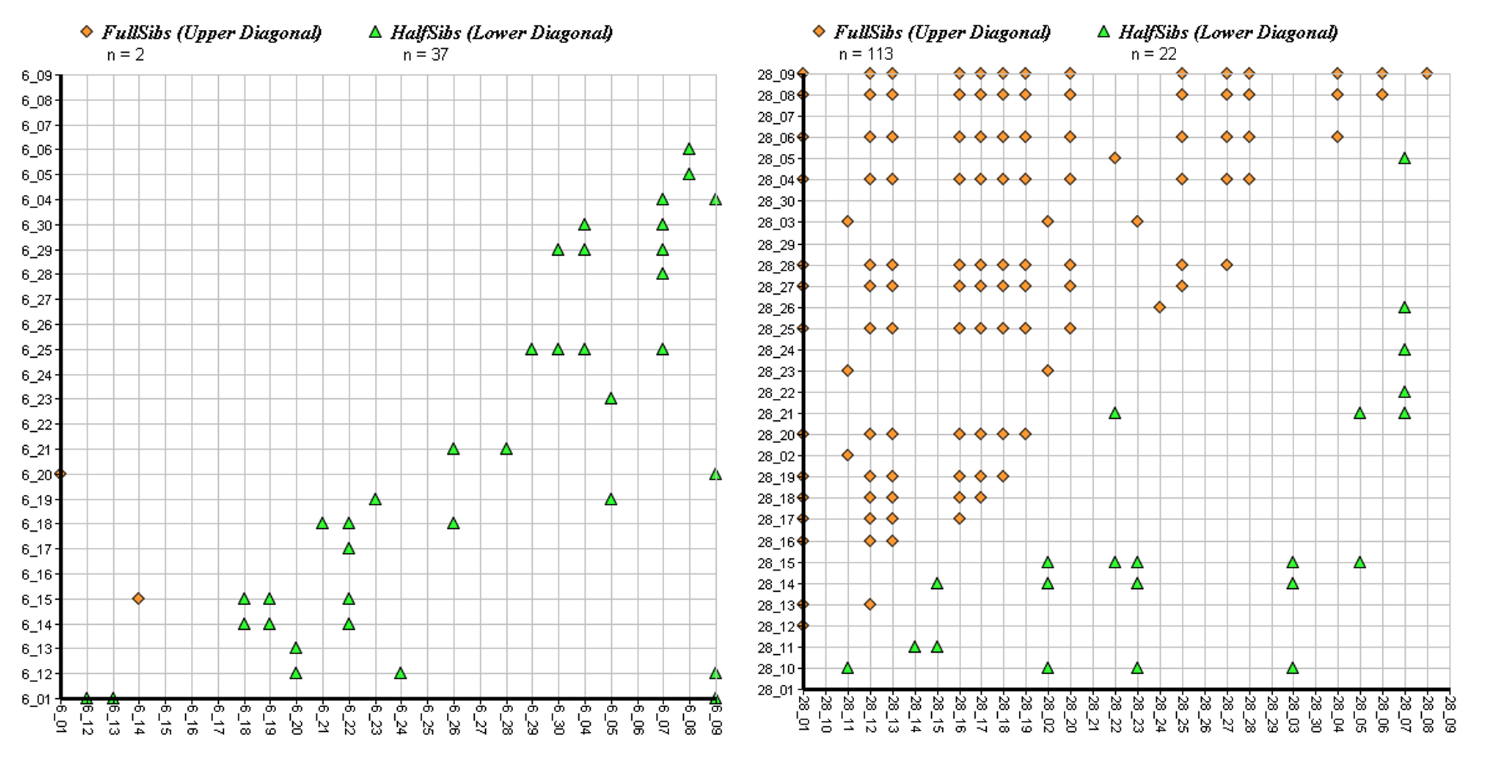
3.4. Sibship Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354, aaf8957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemery, G.E.; Clark, J.R.; Aldinger, E.; Claessens, H.; Malvolti, M.E.; O’connor, E.; Raftoyannis, Y.; Savill, P.S.; Brus, R. Growing scattered broadleaved tree species in Europe in a changing climate: A review of risks and opportunities. Forestry 2010, 83, 65–81. [Google Scholar] [CrossRef] [Green Version]
- Jalonen, R.; Valette, M.; Boshier, D.; Duminil, J.; Thomas, E. Forest and landscape restoration severely constrained by a lack of attention to the quantity and quality of tree seed: Insights from a global survey. Conserv. Lett. 2018, 11, e12424. [Google Scholar] [CrossRef]
- Gömöry, D.; Himanen, K.; Tollefsrud, M.M.; Uggla, C.; Kraigher, H.; Alizoti, P.; Frank, A.; Proschowsky, G.F.; Geburek, T.; Guibert, M.; et al. Genetic Aspects Linked to Production and Use of Forest Reproductive Material (FRM); European Forest Genetic Resources Programme (EUFORGEN); European Forest Institute: Barcelona, Spain, 2021; 216p. [Google Scholar]
- Roloff, A.; Pietzarka, U. Acer platanoides Linne 1753. In Enzyklopädie der Holzgewächse: Handbuch und Atlas der Dendrologie; Wiley-VCH Verlag GmbH & Co.: Weinheim, Germany, 1998; p. 16. [Google Scholar]
- Renner, S.S.; Beenken, L.; Grimm, G.W.; Kocyan, A.; Ricklefs, R.E. The evolution of dioecy, heterodichogamy, and labile sex expression in Acer. Evol. 2007, 61, 2701–2719. [Google Scholar] [CrossRef]
- Kunz, J.; Räder, A.; Bauhus, J. Effects of drought and rewetting on growth and gas exchange of minor European broadleaved tree species. Forests 2016, 7, 239. [Google Scholar] [CrossRef]
- Carón, M.M.; De Frenne, P.; Brunet, J.; Chabrerie, O.; Cousins, S.A.O.; De Backer, L.; Decocq, G.; Diekmann, M.; Heinken, T.; Kolb, A.; et al. Interacting effects of warming and drought on regeneration and early growth of Acer pseudoplatanus and A. platanoides. Plant Biol. 2015, 17, 52–62. [Google Scholar] [CrossRef]
- Lefèvre, F. Human impacts on forest genetic resources in the temperate zone: An updated review. For. Ecol. Manag. 2004, 197, 257–271. [Google Scholar] [CrossRef]
- Lefèvre, F.; Koskela, J.; Hubert, J.; Kraigher, H.; Longauer, R.; Olrik, D.C.; Schüler, S.; Bozzano, M.; Alizoti, P.; Bakys, R.; et al. Dynamic Conservation of Forest Genetic Resources in 33 European Countries. Conserv. Biol. 2013, 27, 373–384. [Google Scholar] [CrossRef]
- Palmberg-Lerche, C.; Turok, J.; Sigaud, P. Forest genetic resources in the international context: Processes, agreements and programmes. In Conservation and Management of Forest Genetic Resources in Europe; Geburek, T., Turok, J., Eds.; Arbora: Zvolen, Slovakia, 2005; pp. 45–73. [Google Scholar]
- Konnert, M.; Fady, B.; Gömöry, D.; A’Hara, S.; Wolter, F.; Ducci, F.; Koskela, J.; Bozzano, M.; Maaten, T.; Kowalczyk, J. Use and Transfer of Forest Reproductive Material: In Europe in the Context of Climate Change; Bioversity International, Ed.; Bioversity International: Rome, Italy, 2015; ISBN 9789292550318. [Google Scholar]
- Streiff, R.; Labbe, T.; Bacilieri, R.; Steinkellner, H.; Glössl, J.; Kremer, A. Within-population genetic structure in Quercus robur L. and Quercus petraea (Matt.) Liebl. assessed with isozymes and microsatellites. Mol. Ecol. 1998, 7, 317–328. [Google Scholar] [CrossRef]
- Hosius, B.; Leinemann, L.; Bergmann, F.; Maurer, W.D.; Tabel, U. Genetische Untersuchungen zu Famielienstrukturen und zur Zwieselbildung in Buchenbeständen. Forst Holz 2003, 58, 51–54. [Google Scholar]
- Leonardi, S.; Raddi, S.; Borghetti, M. Spatial autocorrelation of allozyme traits in a Norway spruce (Picea abies) population. Can. J. For. Res. 1996, 26, 63–71. [Google Scholar] [CrossRef]
- Hosius, B.; Leinemann, L.; Konnert, M.; Bergmann, F. Genetic aspects of forestry in the Central Europe. Eur. J. For. Res. 2006, 125, 407–417. [Google Scholar] [CrossRef]
- Kavaliauskas, D.; Fussi, B.; Westergren, M.; Aravanopoulos, F.; Finzgar, D.; Baier, R.; Alizoti, P.; Bozic, G.; Avramidou, E.; Konnert, M.; et al. The interplay between forest management practices, genetic monitoring, and other long-term monitoring systems. Forests 2018, 9, 133. [Google Scholar] [CrossRef] [Green Version]
- Ziehe, M.; Hattemer, H.H. Auswirkungen räumlicher Verteilungen genetischer Varianten in Buchenbeständen auf dort geerntetes Saatgut. In Mitteilungen aus der Forschungsanstalt für Waldökologie und Forstwirtschaft Rheinland-Pfalz; FAWF: Gwynedd, Wales, 2004; pp. 102–120. [Google Scholar]
- Wojacki, J.; Eusemann, P.; Ahnert, D.; Pakull, B.; Liesebach, H. Genetic diversity in seeds produced in artificial Douglas-fir (Pseudotsuga menziesii) stands of different size. For. Ecol. Manag. 2019, 438, 18–24. [Google Scholar] [CrossRef]
- Westergren, M.; Bozic, G.; Kraigher, H. Trends in forest seed and seedling production in Slovenia. Gozdarski Vestn. 2017, 75, 184–191. [Google Scholar]
- Nickolas, H.; Harrison, P.A.; Tilyard, P.; Vaillancourt, R.E.; Potts, B.M. Inbreeding depression and differential maladaptation shape the fitness trajectory of two co-occurring Eucalyptus species. Ann. For. Sci. 2019, 76, 10. [Google Scholar] [CrossRef] [Green Version]
- Woods, J.H.; Heaman, J.C. Effect of different inbreeding levels on filled seed production in Douglas-fir. Can. J. For. Res. 1989, 19, 54–59. [Google Scholar] [CrossRef]
- Griffin, A.R.; Lindgren, D. Effect of inbreeding on production of filled seed in Pinus radiata—Experimental results and a model of gene action. Theor. Appl. Genet. 1985, 71, 334–343. [Google Scholar] [CrossRef]
- Kormuták, A.; Lindgren, D. Mating system and empty seeds in silver fir (Abies alba mill.). Int. J. For. Genet. 1997, 3, 231–235. [Google Scholar]
- Kärkkäinen, K.; Savolainen, O. The degree of early inbreeding depression determines the selfing rate at the seed stage: Model and results from Pinus sylvestris (scots pine). Heredity 1993, 71, 160–166. [Google Scholar] [CrossRef]
- Mimura, M.; Aitken, S.N. Increased selfing and decreased effective pollen donor number in peripheral relative to central populations in Picea sitchensis (Pinaceae). Am. J. Bot. 2007, 94, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Robledo-Arnuncio, J.J.; Alía, R.; Gil, L. Increased selfing and correlated paternity in a small population of a predominantly outcrossing conifer, Pinus sylvestris. Mol. Ecol. 2004, 13, 2567–2577. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, F.C.; Miles, R.S. Inbreeding depression in height, height growth, and survival of Douglas-fir, ponderosa pine, and noble fir to 10 years of age. For. Sci. 1982, 28, 283–292. [Google Scholar] [CrossRef]
- Doerksen, T.K.; Bousquet, J.; Beaulieu, J. Inbreeding depression in intra-provenance crosses driven by founder relatedness in white spruce. Tree Genet. Genomes 2014, 10, 203–212. [Google Scholar] [CrossRef]
- Lobo, J.A.; Jimenez, D.; Solis-Hernandez, W.; Fuchs, E.J. Lack of early inbreeding depression and distribution of selfing rates in the neotropical emergent tree Ceiba pentandra: Assessment from several reproductive events. Am. J. Bot. 2015, 102, 983–991. [Google Scholar] [CrossRef] [Green Version]
- Crnokrak, P.; Barrett, S.C.H. Perspective: Purging the genetic load: A review of the experimental evidence. Evolution 2002, 56, 2347–2358. [Google Scholar] [CrossRef]
- Charlesworth, D.; Willis, J.H. The genetics of inbreeding depression. Nat. Rev. Genet. 2009, 10, 783–796. [Google Scholar] [CrossRef]
- Losdat, S.; Chang, S.M.; Reid, J.M. Inbreeding depression in male gametic performance. J. Evol. Biol. 2014, 27, 992–1011. [Google Scholar] [CrossRef]
- Aguilar, R.; Cristóbal-Pérez, E.J.; Balvino-Olvera, F.J.; de Jesús Aguilar-Aguilar, M.; Aguirre-Acosta, N.; Ashworth, L.; Lobo, J.A.; Martén-Rodríguez, S.; Fuchs, E.J.; Sanchez-Montoya, G.; et al. Habitat fragmentation reduces plant progeny quality: A global synthesis. Ecol. Lett. 2019, 22, 1163–1173. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, R.; Ashworth, L.; Galetto, L.; Aizen, M.A. Plant reproductive susceptibility to habitat fragmentation: Review and synthesis through a meta-analysis. Ecol. Lett. 2006, 9, 968–980. [Google Scholar] [CrossRef]
- Aguilar, R.; Quesada, M.; Ashworth, L.; Herrerias-Diego, Y.; Lobo, J. Genetic consequences of habitat fragmentation in plant populations: Susceptible signals in plant traits and methodological approaches. Mol. Ecol. 2008, 17, 5177–5188. [Google Scholar] [CrossRef] [PubMed]
- Young, A.; Boyle, T.; Brown, T. The population genetic consequences of habitat fragmentation for plants. Trends Ecol. Evol. 1996, 11, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Hamrick, J.L.; Godt, M.J.W.; Sherman-Broyles, S.L. Factors influencing levels of genetic diversity in woody plant species. New For. 1992, 6, 95–124. [Google Scholar] [CrossRef]
- Hamrick, J.L. Response of forest trees to global environmental changes. For. Ecol. Manag. 2004, 197, 323–335. [Google Scholar] [CrossRef]
- Vranckx, G.; Jacquemyn, H.; Muys, B.; Honnay, O. Meta-Analysis of Susceptibility of Woody Plants to Loss of Genetic Diversity through Habitat Fragmentation. Conserv. Biol. 2012, 26, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Hipp, A.L.; Manos, P.S.; González-Rodríguez, A.; Hahn, M.; Kaproth, M.; McVay, J.D.; Avalos, S.V.; Cavender-Bares, J. Sympatric parallel diversification of major oak clades in the Americas and the origins of Mexican species diversity. New Phytol. 2018, 217, 439–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bittkau, C. Charakterisierung der Genetischen Variation Europäischer Populationen von Acer spp. und Populus tremula auf der Basis der Chloroplaten-DNA: Rückschlüsse auf die Postglaziale Ausbreitung und Differenzierung Forstlicher Provenienzen; FAWF: Gwynedd, Wales, 2002; pp. 1–87. [Google Scholar]
- Neophytou, C.; Konnert, M.; Fussi, B. Western and eastern post-glacial migration pathways shape the genetic structure of sycamore maple (Acer pseudoplatanus L.) in Germany. For. Ecol. Manag. 2019, 432, 83–93. [Google Scholar] [CrossRef]
- Liepelt, S.; Bialozyt, R.; Ziegenhagen, B. Wind-dispersed pollen mediates postglacial gene flow among refugia. Proc. Natl. Acad. Sci. USA. 2002, 99, 14590–14594. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, E. Der Ahorn. Wald-, Park- und Straßenbaum; VEB Deutscher Landwirtschaftsverlag: Leipzig, Germany, 1960. [Google Scholar]
- Rusanen, M.; Vakkari, P.; Blom, A. Evaluation of the finnish gene-conservation strategy for Norway maple (Acer platanoides L.) in the light of allozyme variation. For. Genet. 2000, 7, 155–165. [Google Scholar]
- Rusanen, M.; Vakkari, P.; Blom, A. Genetic structure of Acer platanoides and Betula pendula in northern Europe. Can. J. For. Res. 2003, 33, 1110–1115. [Google Scholar] [CrossRef]
- Perry, D.J.; Knowles, P. Allozyme variation in sugar maple at the northern limit of its range in Ontario, Canada. Can. J. For. Res. 1989, 19, 509–514. [Google Scholar] [CrossRef]
- Fore, S.A.; Hickey, R.J.; Vankat, J.L.; Guttman, S.I.; Schaefer, R.L. Genetic structure after forest fragmentation: A landscape ecology perspective. Can. J. Bot. 1992, 70, 1659–1668. [Google Scholar] [CrossRef]
- Koskela, J.; Lefèvre, F.; Schueler, S.; Kraigher, H.; Olrik, D.C.; Hubert, J.; Longauer, R.; Bozzano, M.; Yrjänä, L.; Alizoti, P.; et al. Translating conservation genetics into management: Pan-European minimum requirements for dynamic conservation units of forest tree genetic diversity. Biol. Conserv. 2013, 157, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Kavaliauskas, D.; Šeho, M.; Baier, R.; Fussi, B. Genetic variability to assist in the delineation of provenance regions and selection of seed stands and gene conservation units of wild service tree (Sorbus torminalis (L.) Crantz) in southern Germany. Eur. J. For. Res. 2021, 140, 551–565. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data [Online]. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 1 March 2022).
- Rousset, F. GENEPOP’007: A complete re-implementation of the GENEPOP software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Szpiech, Z.A.; Jakobsson, M.; Rosenberg, N.A. ADZE: A rarefaction approach for counting alleles private to combinations of populations. Bioinformatics 2008, 24, 2498–2504. [Google Scholar] [CrossRef] [Green Version]
- Goudet, J. FSTAT (Version 1.2): A Computer Program to Calculate F-Statistics. J. Hered. 1995, 86, 485–486. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. PHYLIP (Phylogeny Inference Package); Department of Genome Sciences, University of Washington: Seattle, WA, USA, 2005. [Google Scholar]
- Piry, S.; Luikart, G.; Cornuet, J.M. BOTTLENECK: A computer program for detecting recent reductions in the effective population size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Queller, D.C.; Goodnight, K.F. Estimating Relatedness Using Genetic Markers Published by: Society for the Study of Evolution Stable. Evolution (N. Y). 2016, 43, 258–275. [Google Scholar]
- Lynch, M.; Ritland, K. Estimation of pairwise relatedness with molecular markers. Genetics 1999, 152, 1753–1766. [Google Scholar] [CrossRef] [PubMed]
- Jones, O.R.; Wang, J. COLONY: A program for parentage and sibship inference from multilocus genotype data. Mol. Ecol. Resour. 2010, 10, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Ruibal, M.; Lindenmayer, D.B. Spatial autocorrelation analysis offers new insights into gene flow in the Australian bush rat, Rattus fuscipes. Evolution 2003, 57, 1182–1195. [Google Scholar] [CrossRef]
- Moran, P. Notes on Continuous Stochastic Phenomena. Biometrika 1950, 37, 17–23. Available online: http://www.jstor.org/stable/2332142 (accessed on 1 March 2022). [CrossRef]
- Savolainen, O.; Pyhäjärvi, T.; Knürr, T. Gene flow and local adaptation in trees. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 595–619. [Google Scholar] [CrossRef]
- Blanc-Jolivet, C.; Degen, B. Using simulations to optimize genetic diversity in Prunus avium seed harvests. Tree Genet. Genomes 2014, 10, 503–512. [Google Scholar] [CrossRef]
- Guarino, C.; Santoro, S.; De Simone, L.; Cipriani, G. Prunus avium: Nuclear DNA study in wild populations and sweet cherry cultivars. Genome 2009, 52, 320–337. [Google Scholar] [CrossRef] [PubMed]
- George, J.P.; Konrad, H.; Collin, E.; Thevenet, J.; Ballian, D.; Idzojtic, M.; Kamm, U.; Zhelev, P.; Geburek, T. High molecular diversity in the true service tree (Sorbus domestica) despite rareness: Data from Europe with special reference to the Austrian occurrence. Ann. Bot. 2015, 115, 1105–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobeš, C.; Konrad, H.; Geburek, T. Potential population genetic consequences of habitat fragmentation in central European forest trees and associated understorey species-An introductory survey. Diversity 2017, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Akhmetov, A.; Ianbaev, R.; Boronnikova, S.; Yanbaev, Y.; Gabitova, A.; Kulagin, A. Norway maple (Acer platanoides) and pedunculate oak (Quercus robur) demonstrate different patterns of genetic variation within and among populations on the eastern border of distribution ranges. J. For. Sci. 2021, 67, 522–532. [Google Scholar] [CrossRef]
- Firbas, F. Spät-und Nacheiszeitliche Waldgeschichte Mitteleuropas Nördlich der Alpen; Fischer: Jena, Germany, 1949. [Google Scholar]
- Bertsch, K. Geschichte des Deutschen Waldes; G. Fischer: Jena, Germany, 1949. [Google Scholar]
- Tollefsrud, M.M.; Sønstebø, J.H.; Brochmann, C.; Johnsen, Ø.; Skrøppa, T.; Vendramin, G.G. Combined analysis of nuclear and mitochondrial markers provide new insight into the genetic structure of North European Picea abies. Heredity 2009, 102, 549–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caudullo, G.; de Rigo, D. Acer platanoides in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; Publications Office of the EU: Luxembourg, Germany, 2016; p. e019159. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population Number | Name | Country | Province | Latitude | Longitude | Number of Samples | Comments |
|---|---|---|---|---|---|---|---|
| 1 | Bromatsreute | AT | Vorarlberg | 47.54 | 9.82 | 29 | Nature reserve forest |
| 2 | Zwurms | AT | Vorarlberg | 47.33 | 9.66 | 30 | Natural |
| 3 | Schruns | AT | Vorarlberg | 47.08 | 9.90 | 17 | Natural |
| 4 | Bezau | AT | Vorarlberg | 47.39 | 9.86 | 29 | Natural |
| 5 | Zams | AT | Tyrol | 47.16 | 10.60 | 20 | Natural |
| 6 | Mühlgraben | AT | Lower Austria | 48.03 | 15.55 | 26 | Nature reserve forest |
| 7 | Kollmitzgraben | AT | Lower Austria | 48.83 | 15.54 | 30 | Nature reserve forest |
| 8 | Mitterberg | AT | Upper Austria | 48.43 | 13.57 | 29 | Nature reserve forest |
| 9 | Stauf | AT | Upper Austria | 48.44 | 13.92 | 45 | Nature reserve forest |
| 10 | Marsbach | AT | Upper Austria | 48.46 | 13.82 | 20 | Nature reserve forest |
| 11 | FreyensteinerDonauwald | AT | Lower Austria | 48.23 | 14.93 | 30 | Nature reserve forest |
| 12 | Dobra | AT | Lower Austria | 48.59 | 15.40 | 30 | Nature reserve forest |
| 13 | Kardinalgraben | AT | Lower Austria | 48.11 | 15.23 | 40 | Nature reserve forest |
| 14 | Luxensteinwald | AT | Lower Austria | 48.64 | 14.74 | 32 | Nature reserve forest |
| 15 | Sommerein | AT | Lower Austria | 47.99 | 16.68 | 20 | Nature reserve forest |
| 16 | Sankt Gertraud | AT | Carinthia | 46.52 | 14.60 | 34 | Natural |
| 17 | Leobersdorf | AT | Lower Austria | 47.92 | 16.13 | 28 | Natural |
| 18 | Landsberg | AT | Upper Austria | 47.90 | 14.26 | 30 | Natural |
| 19 | Salzkammergut | AT | Salzburg/Styria | 47.74 | 13.42 | 30 | Natural |
| 20 | Tennengau | AT | Salzburg | 47.65 | 13.10 | 30 | Natural |
| 21 | Vorderberg | AT | Carinthia | 46.58 | 13.55 | 24 | Natural |
| 22 | Schönbrunn | AT | Vienna | 48.18 | 16.30 | 29 | Natural |
| 23 | Seebenstein | AT | Lower Austria | 47.63 | 16.13 | 38 | Natural |
| 24 | Falkenstein | AT | Lower Austria | 48.73 | 16.56 | 18 | Natural |
| 25 | Klagenfurt | AT | Carinthia | 46.67 | 14.31 | 20 | Planted stand |
| 26 | Grünbach | AT | Lower Austria | 47.80 | 15.99 | 33 | natural |
| 27 | Pernitz | AT | Lower Austria | 47.93 | 15.98 | 11 | natural |
| 28 | Bernhardtsthal | AT | Lower Austria | 48.70 | 16.90 | 30 | Registered Seed Stand/planted |
| 29 | Planting stock Austria | AT | 24 | Obtained from nursery as putative offspring from Bernhardtsthal (pop. 28) | |||
| 30 | Sternberg | AT | Carinthia | 46.63 | 13.98 | 24 | Semi-urban, isolated population |
| 31 | Germany1 | DE | Baden-Würtemberg | 48.60 | 9.01 | 25 | Obtained from nursery |
| 32 | Germany2 | DE | Baden-Würtemberg | 48.53 | 8.99 | 36 | Obtained from nursery |
| 33 | Borjád | HU | Borjád | 45.93 | 18.48 | 25 | Obtained from nursery |
| 34 | Szombathely | HU | Szombathely | 47.20 | 16.55 | 26 | Obtained from nursery |
| 35 | Busovaca | BH | Busovaca | 44.10 | 17.90 | 31 | Natural Forest |
| 36 | Radava | BH | Radava | 43.89 | 18.42 | 30 | Natural Forest |
| 37 | Estonia | EE | Viljandimaa | 58.09 | 25.06 | 14 | Natural Forest |
| 38 | Haarikonmäki | FI | East Finland | 61.57 | 29.18 | 31 | Gene conservation unit |
| Locus | Size bp | Mean Na | Repeat Motif | Primer-F | Primer-R | GenBank Accession No. | Null Alleles |
|---|---|---|---|---|---|---|---|
| Ap_20237 | 179–203 | 8.895 | (TATT)8 | AGTTACCTCTCAAAATCTCCATCTAAC | AATAAGGGGGTGGTGTGGAC | MT895404 | 0.105 |
| Ap_188503 | 245–258 | 5.368 | (TTTA)7 | TCGTTCCTTATCGGGCAAAATG | TCAGAATTGTTTTCTTCACTAGTTGC | MT895405 | 0.051 |
| Ap_541809 | 112–136 | 4.000 | (TTA)10 | TGAGAATTTTCCAAGTACTAGAACCG | TCAAGCACTAGAACTATTTATCCCG | MT895406 | 0.016 |
| Ap_638034 | 175–196 | 10.553 | (ACAT)7 | GAGATTGCAAGGCCCTGTTG | ATCATCTGTTGAGCCCCTCC | MT895407 | 0.019 |
| Ap_33364 | 228–239 | 7.316 | (TTTA)7 | CGGTGGAAACCGACTGTAATC | TCATAATGTTTGGAAGGTTTTGGC | MT895408 | 0.022 |
| Ap_44827 | 92–105 | 8.684 | (TA)18 | GGAGCATCACGAAACTACCG | AAATGGCGTCACTTGCTACG | MT895409 | 0.017 |
| Ap_64069 | 85–106 | 6.868 | (ATA)14 | ACATTAAAAGTGGAAACTGAAAAGG | ACCGTCGTTTAGATATAGACATGC | MT895410 | 0.027 |
| Ap_274758 | 67–183 | 10.026 | (AG)15 | GGTATTAACAGAATTAGGAAGGGGG | AGTGTCACAAAGTTCGCCAG | MT895411 | 0.017 |
| Ap_361727 | 186–204 | 4.579 | (ATA)9 | GCAACTCCAAATCCGGACAG | CGGTTAGCTGTCATCGTTTCTC | MT895412 | 0.093 |
| Ap_374910 | 224–252 | 7.921 | (ATT)15 | ACAAGTGGAGTCGTATAGATCCTG | ACTTCGAGTTCTGATTAAAATTGGG | MT895413 | 0.072 |
| Ap_411647 | 227–245 | 9.053 | (AT)19 | TCGGCTATTCATCTCAAGATTTGG | TGGGTAACCAATCGGTCCAG | MT895414 | 0.057 |
| Ap_488207 | 89–182 | 3.711 | (AAAT)7 | TCGGGAAACTGGTTAGGTGC | GCAACTTTGGAGCGGTATCC | MT895415 | 0.134 |
| Ap_501482 | 233–241 | 5.658 | (TA)11 | AGGCTTCCATGGCGAAATTG | CTT-GCTTGACCAGAGGTTTCC | MT895416 | 0.007 |
| Ap_658195 | 190–206 | 3.974 | (TACA)8 | AGGTCTAGGACCCAGGAAATC | GGTTTAGCTGGTTGAGATGGTC | MT895417 | 0.105 |
| Ap_671221 | 123–214 | 6.895 | (TATT)7 | TGGAATGTAAAGAGACATTAGTGG | GTGGCTCCATTACCACCATC | MT895418 | 0.018 |
| Pop. | Ne | Ho | He | uHe | F (Sign. HZ Deficit/Excess) | Ar | Pop_eff (CI) | Bottleneck IAM | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 3.534 | ± | 0.312 | 0.670 | ± | 0.035 | 0.694 | ± | 0.027 | 0.706 | ± | 0.028 | 0.033 | ± | 0.036 | * | 4.764 | ± | 0.337 | 38 | 22–66 | 0.0257 | ± | 0.0065 |
| 2 | 3.847 | ± | 0.305 | 0.745 | ± | 0.036 | 0.724 | ± | 0.021 | 0.736 | ± | 0.021 | −0.029 | ± | 0.038 | 4.851 | ± | 0.385 | 34 | 21–59 | 0.1004 | ± | 0.0013 | |
| 3 | 3.648 | ± | 0.425 | 0.661 | ± | 0.054 | 0.686 | ± | 0.038 | 0.707 | ± | 0.039 | 0.043 | ± | 0.046 | 4.981 | ± | 0.460 | 26 | 14–53 | 0.1004 | ± | 0.0383 | |
| 4 | 3.657 | ± | 0.308 | 0.717 | ± | 0.029 | 0.707 | ± | 0.024 | 0.719 | ± | 0.025 | −0.020 | ± | 0.041 | * | 4.901 | ± | 0.396 | 45 | 27–80 | 0.0236 | ± | 0.0062 |
| 5 | 3.641 | ± | 0.418 | 0.613 | ± | 0.031 | 0.698 | ± | 0.026 | 0.717 | ± | 0.026 | 0.115 | ± | 0.050 | *** | 4.635 | ± | 0.442 | 30 | 17–65 | 0.0028 | ± | 0.0036 |
| 6 | 4.206 | ± | 0.355 | 0.727 | ± | 0.061 | 0.742 | ± | 0.025 | 0.756 | ± | 0.025 | 0.032 | ± | 0.073 | 5.256 | ± | 0.414 | 28 | 15–54 | 0.0029 | ± | 0.0003 | |
| 7 | 3.939 | ± | 0.390 | 0.724 | ± | 0.051 | 0.715 | ± | 0.033 | 0.727 | ± | 0.034 | −0.008 | ± | 0.043 | 5.135 | ± | 0.426 | 48 | 29–84 | 0.0027 | ± | 0.0024 | |
| 8 | 4.534 | ± | 0.611 | 0.674 | ± | 0.054 | 0.724 | ± | 0.046 | 0.736 | ± | 0.046 | 0.060 | ± | 0.058 | *** | 5.596 | ± | 0.576 | 85 | 51–165 | 0.0272 | ± | 0.0079 |
| 9 | 4.293 | ± | 0.453 | 0.739 | ± | 0.042 | 0.734 | ± | 0.032 | 0.742 | ± | 0.033 | −0.002 | ± | 0.021 | 5.411 | ± | 0.475 | 51 | 33–81 | 0.0027 | ± | 0.0009 | |
| 10 | 4.051 | ± | 0.423 | 0.661 | ± | 0.048 | 0.728 | ± | 0.026 | 0.746 | ± | 0.026 | 0.095 | ± | 0.050 | ** | 5.461 | ± | 0.468 | 54 | 31–122 | 0.0283 | ± | 0.0184 |
| 11 | 3.672 | ± | 0.392 | 0.603 | ± | 0.060 | 0.672 | ± | 0.061 | 0.683 | ± | 0.062 | 0.092 | ± | 0.035 | *** | 4.825 | ± | 0.446 | 36 | 22–65 | 0.0240 | ± | 0.0197 |
| 12 | 4.460 | ± | 0.524 | 0.730 | ± | 0.046 | 0.737 | ± | 0.035 | 0.750 | ± | 0.035 | 0.014 | ± | 0.029 | * | 5.531 | ± | 0.487 | 44 | 26–82 | 0.0032 | ± | 0.0016 |
| 13 | 4.634 | ± | 0.633 | 0.770 | ± | 0.036 | 0.742 | ± | 0.033 | 0.751 | ± | 0.033 | −0.041 | ± | 0.025 | 5.533 | ± | 0.522 | 40 | 26–65 | 0.0031 | ± | 0.0003 | |
| 14 | 4.136 | ± | 0.545 | 0.670 | ± | 0.065 | 0.698 | ± | 0.054 | 0.709 | ± | 0.055 | 0.037 | ± | 0.052 | * | 4.995 | ± | 0.510 | 39 | 23–68 | 0.0209 | ± | 0.0009 |
| 15 | 4.701 | ± | 0.593 | 0.791 | ± | 0.048 | 0.745 | ± | 0.036 | 0.764 | ± | 0.037 | −0.057 | ± | 0.029 | 5.772 | ± | 0.569 | 33 | 18–68 | 0.0032 | ± | 0.0005 | |
| 16 | 4.370 | ± | 0.538 | 0.729 | ± | 0.041 | 0.738 | ± | 0.029 | 0.749 | ± | 0.029 | 0.017 | ± | 0.025 | 5.299 | ± | 0.485 | 29 | 18–53 | 0.0027 | ± | 0.0003 | |
| 17 | 3.886 | ± | 0.427 | 0.711 | ± | 0.067 | 0.688 | ± | 0.057 | 0.701 | ± | 0.058 | −0.005 | ± | 0.043 | 5.150 | ± | 0.545 | 36 | 22–63 | 0.1009 | ± | 0.0222 | |
| 18 | 5.092 | ± | 0.698 | 0.793 | ± | 0.038 | 0.760 | ± | 0.035 | 0.773 | ± | 0.035 | −0.047 | ± | 0.027 | 5.898 | ± | 0.628 | 58 | 36–100 | 0.0030 | ± | 0.0002 | |
| 19 | 4.262 | ± | 0.464 | 0.765 | ± | 0.041 | 0.733 | ± | 0.033 | 0.745 | ± | 0.034 | −0.040 | ± | 0.022 | 5.330 | ± | 0.481 | 51 | 33–88 | 0.1007 | ± | 0.0014 | |
| 20 | 4.330 | ± | 0.533 | 0.724 | ± | 0.036 | 0.736 | ± | 0.029 | 0.749 | ± | 0.030 | 0.017 | ± | 0.030 | * | 5.528 | ± | 0.498 | 64 | 40–116 | 0.0270 | ± | 0.0032 |
| 21 | 3.647 | ± | 0.404 | 0.632 | ± | 0.059 | 0.681 | ± | 0.043 | 0.696 | ± | 0.044 | 0.070 | ± | 0.067 | ** | 5.053 | ± | 0.505 | 35 | 20–69 | 0.1018 | ± | 0.0228 |
| 22 | 4.605 | ± | 0.558 | 0.765 | ± | 0.045 | 0.744 | ± | 0.036 | 0.757 | ± | 0.037 | −0.026 | ± | 0.028 | 5.675 | ± | 0.515 | 46 | 28–82 | 0.0281 | ± | 0.0031 | |
| 23 | 4.376 | ± | 0.471 | 0.736 | ± | 0.045 | 0.741 | ± | 0.029 | 0.751 | ± | 0.030 | 0.012 | ± | 0.043 | * | 5.540 | ± | 0.492 | 48 | 32–80 | 0.0029 | ± | 0.0012 |
| 24 | 3.991 | ± | 0.543 | 0.702 | ± | 0.049 | 0.705 | ± | 0.035 | 0.725 | ± | 0.036 | 0.013 | ± | 0.041 | 5.275 | ± | 0.615 | 38 | 19–91 | 0.2424 | ± | 0.0129 | |
| 25 | 4.186 | ± | 0.500 | 0.651 | ± | 0.050 | 0.720 | ± | 0.038 | 0.739 | ± | 0.039 | 0.091 | ± | 0.055 | *** | 5.183 | ± | 0.466 | 54 | 30–151 | 0.0293 | ± | 0.0022 |
| 26 | 4.769 | ± | 0.669 | 0.749 | ± | 0.033 | 0.750 | ± | 0.031 | 0.762 | ± | 0.032 | 0.000 | ± | 0.021 | 5.643 | ± | 0.560 | 53 | 33–92 | 0.0274 | ± | 0.0017 | |
| 27 | 4.154 | ± | 0.513 | 0.777 | ± | 0.048 | 0.719 | ± | 0.034 | 0.754 | ± | 0.036 | −0.075 | ± | 0.035 | 5.705 | ± | 0.687 | 55 | 24–undef | 0.2784 | ± | 0.0298 | |
| 28 | 3.087 | ± | 0.327 | 0.742 | ± | 0.060 | 0.636 | ± | 0.040 | 0.647 | ± | 0.040 | −0.159 | ± | 0.052 | *** | 4.250 | ± | 0.390 | 7 | 4–21 | 0.0199 | ± | 0.0366 |
| 29 | 2.757 | ± | 0.423 | 0.598 | ± | 0.092 | 0.544 | ± | 0.065 | 0.556 | ± | 0.067 | −0.091 | ± | 0.092 | * | 3.598 | ± | 0.443 | 17 | 8–38 | 0.0576 | ± | 0.0290 |
| 30 | 2.963 | ± | 0.216 | 0.686 | ± | 0.048 | 0.638 | ± | 0.036 | 0.652 | ± | 0.037 | −0.065 | ± | 0.043 | 3.933 | ± | 0.209 | 21 | 12–43 | 0.0226 | ± | 0.0045 | |
| 31 | 4.636 | ± | 0.597 | 0.663 | ± | 0.059 | 0.739 | ± | 0.038 | 0.755 | ± | 0.039 | 0.124 | ± | 0.052 | *** | 5.398 | ± | 0.534 | 16 | 9–34 | 0.0255 | ± | 0.0005 |
| 32 | 4.606 | ± | 0.498 | 0.730 | ± | 0.028 | 0.754 | ± | 0.028 | 0.765 | ± | 0.028 | 0.029 | ± | 0.025 | * | 5.502 | ± | 0.461 | 32 | 20–57 | 0.0029 | ± | 0.0001 |
| 33 | 4.434 | ± | 0.547 | 0.724 | ± | 0.052 | 0.729 | ± | 0.038 | 0.744 | ± | 0.039 | 0.015 | ± | 0.032 | 5.367 | ± | 0.550 | 27 | 15–51 | 0.0031 | ± | 0.0012 | |
| 34 | 3.863 | ± | 0.402 | 0.675 | ± | 0.058 | 0.706 | ± | 0.035 | 0.720 | ± | 0.036 | 0.051 | ± | 0.056 | * | 5.096 | ± | 0.488 | 14 | 8–30 | 0.0026 | ± | 0.0033 |
| 35 | 3.483 | ± | 0.317 | 0.680 | ± | 0.041 | 0.688 | ± | 0.029 | 0.699 | ± | 0.029 | 0.012 | ± | 0.043 | 4.873 | ± | 0.367 | 46 | 27–82 | 0.0030 | ± | 0.0441 | |
| 36 | 4.780 | ± | 0.741 | 0.718 | ± | 0.055 | 0.743 | ± | 0.034 | 0.756 | ± | 0.034 | 0.036 | ± | 0.056 | * | 5.682 | ± | 0.622 | 42 | 25–77 | 0.0032 | ± | 0.0016 |
| 37 | 3.433 | ± | 0.316 | 0.652 | ± | 0.086 | 0.672 | ± | 0.042 | 0.697 | ± | 0.043 | 0.073 | ± | 0.102 | 4.730 | ± | 0.437 | 52 | 22–1651 | 0.0850 | ± | 0.0167 | |
| 38 | 2.753 | ± | 0.277 | 0.633 | ± | 0.057 | 0.588 | ± | 0.050 | 0.598 | ± | 0.051 | −0.086 | ± | 0.047 | 3.760 | ± | 0.375 | 16 | 9–34 | 0.4203 | ± | 0.0532 | |
| Total | 4.037 | ± | 0.079 | 0.703 | ± | 0.008 | 0.708 | ± | 0.006 | 0.722 | ± | 0.006 | 0.009 | ± | 0.008 | 5.135 | ± | 0.481 | 39.2 | 0.0257 | ± | 0.0065 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazic, D.; George, J.-P.; Rusanen, M.; Ballian, D.; Pfattner, S.; Konrad, H. Population Differentiation in Acer platanoides L. at the Regional Scale—Laying the Basis for Effective Conservation of Its Genetic Resources in Austria. Forests 2022, 13, 552. https://doi.org/10.3390/f13040552
Lazic D, George J-P, Rusanen M, Ballian D, Pfattner S, Konrad H. Population Differentiation in Acer platanoides L. at the Regional Scale—Laying the Basis for Effective Conservation of Its Genetic Resources in Austria. Forests. 2022; 13(4):552. https://doi.org/10.3390/f13040552
Chicago/Turabian StyleLazic, Desanka, Jan-Peter George, Mari Rusanen, Dalibor Ballian, Stefanie Pfattner, and Heino Konrad. 2022. "Population Differentiation in Acer platanoides L. at the Regional Scale—Laying the Basis for Effective Conservation of Its Genetic Resources in Austria" Forests 13, no. 4: 552. https://doi.org/10.3390/f13040552