
Abstract
Purpose of Review
Recognizing that in the context of global change, tree genetic diversity represents a crucial resource for future forest adaptation, we review and highlight the major forest genetics research achievements of the past decades in biodiversity-rich countries of the Mediterranean region. For this, we conducted a bibliometric analysis of the scientific literature spanning the past thirty years (1991–2020). Putting together the representative regionwide expertise of our co-authorship, we propose research perspectives for the next decade.
Recent Findings
Forest genetics research in Mediterranean countries is organized into three different scientific domains of unequal importance. The domain “Population diversity and Differentiation” related to over 62% of all publications of the period, the domain “Environmental conditions, growth and stress response” to almost 23%, and the domain “Phylogeography” to almost 15%. Citation rate was trending the opposite way, indicating a strong and sustained interest in phylogeography and a rising interest for genetics research related to climate change and drought resistance. The share of publications from Asia and Africa to the total within the Mediterranean increased significantly during the 30-year period analyzed, reaching just below 30% during the last decade.
Summary
Describing poorly known species and populations, including marginal populations, using the full potential of genomic methods, testing adaptation in common gardens, and modeling adaptive capacity to build reliable scenarios for forest management remain strategic research priorities. Delineating areas of high and low genetic diversity, for conservation and restoration, respectively, is needed. Joining forces between forest management and forest research, sharing data, experience, and knowledge within and among countries will have to progress significantly, e.g., to assess the potential of Mediterranean genetic resources as assisted migration material worldwide.
Introductory quote:
Let us collect with care the facts we can observe, let us consult experience wherever we can, and when this experience is inaccessible to us, let us assemble all the inductions which observation of facts analogous to those which escape us can furnish and let us assert nothing categorically; in this way, we shall be able little by little to discover the causes of a multitude of natural phenomena, and, perhaps, even of phenomena which seem the most incomprehensible... J.B. de Lamarck (Philosophie zoologique, 1809), cited by O. Langlet (1971).
Similar content being viewed by others


Introduction
The Convention on Biological Diversity (CBD) recognizes that biological diversity, at all levels of organization, from genes to ecosystems, has an intrinsic value and sustains the life systems of the biosphere. As such, biological diversity needs to be protected and its components to be sustainably used (article 1 of the CBD). Its protection and restoration also strongly interact with climate change mitigation and adaptation: “limiting global warming to ensure a habitable climate and protecting biodiversity are mutually supporting goals, and their achievement is essential for sustainably and equitably providing benefits to people” [1••].
However, the decade 2011–2020 failed in halting biodiversity loss worldwide [2••]. Despite significant efforts, most national commitments have fallen short of reaching agreed international targets and only six of the twenty Aichi targets have been partially achieved [3]. In addition to long-term effects, such as land use change for agriculture and urbanization, pollution, and invasive species, climate change is now increasingly recognized as a major driver of biodiversity loss [4, 5]. Therefore, scientists now call for integrated policies and actions to counter climate change and biodiversity loss, particularly in biodiversity rich regions [6, 7].
The Mediterranean basin, a crossroads for the European, Saharan, and Irano-Turanian floristic regions, is one of the 36 global biodiversity hotspots [8, 9]. Its plant diversity is one of the richest in the world. While occupying 1.6% of the Earth’s terrestrial surface, it hosts approximately 7% of the World’s plant taxa [10, 11]. Moreover, almost 25,000 species are native to the Mediterranean basin, more than half being regional endemics [12]. The European Union recognizes the value of this diversity. Out of the 233 habitats of community importance listed in Annex 1 of Habitat Directive 92/43 of the European Union, 117 are occurring in Mediterranean Europe, among which 93 are exclusively found there. This high rate of endemism and habitat diversity is a striking feature of the Mediterranean flora [13, 14], making the Mediterranean basin a key area for the long-term conservation of genetic and taxonomic diversity [15].
The tree flora of the Mediterranean basin is also remarkable for its diversity and its ecology. It is composed of 74 genera. A significant subset of 64 occur in Mediterranean Europe, with 210 tree species, 30 of which being endemics, often narrowly restricted [16••]. The geologic and climatic events of the Tertiary, Pleistocene, and Holocene are responsible for this wealth of diversity in modern Mediterranean forests, despite (and sometimes because of) significant human impact [15, 17, 18••, 19, 20, 21]. The resulting geomorphologic, topographic, and geographic complexity of the region also led to significantly diverse genetic lineages [22, 23, 24].
Mediterranean forests and the biodiversity they shelter have adapted to unique climatic factors, such as pronounced bi-seasonality characterized by drought episodes unfavorable to growth during the warm summer season and large year to year variation in total rainfall [25]. They have also endured millennia-long human impact, mainly fragmentation and overexploitation for multiple goods and services, including land use change for agriculture and grazing [13, 26].
There were an estimated 88 million hectares (ha) of forest area in Mediterranean countries in 2015, representing 2.2% of the world’s total forest area of 4.06 billion ha [2••]. Forests of Southern Europe represent 73% of the Mediterranean total, while 18% are in west Asia and 9% in North Africa [27]. Adopting a more restricted bioclimatic point of view, Mediterranean type forests span circa 25.5 million ha, with an additional circa 50 million ha of scrubland and open forests that are common vegetation types in the Mediterranean [28]. These formations have expanded at a net rate of 0.85% per year between 1990 and 2010, mostly as a result of natural expansion (0.67% per year), with reforestation contributing 0.23% per year to this expansion. Deforestation remains at the low level of 0.05% per year, although it is trending upwards [28].
Despite an overall expansion, over 5 million hectares of forests were lost during the 2000–2020 period, of which the vast majority occurred during the second decade of the period. Almost two-thirds of forest cover loss occurred in southwestern Europe, and was mainly due to windstorms and wildfires, possibly aggravated by climate change, but also due to forestry activities. Land clearing for agriculture and urbanization was a major driver of forest loss in southern and eastern Mediterranean countries (Global Forest Watch, https://www.globalforestwatch.org/, accessed June 2021). Forest degradation where forest is overexploited as cattle range or for fuelwood, and increasing risks where forests are abandoned, are important sources of concern in the region [28]. Climate change is now the most recent of human impacts of significance for Mediterranean forests [29]. In an era of emerging “mega-disturbances” [30] when several main drivers of change are acting simultaneously and disturbance is increased by climate change, the hyper-diverse Mediterranean Basin is particularly vulnerable and at risk of biodiversity loss [29, 31, 32, 33], especially its forests, in spite of their current expansion [26, 28].
Genetic diversity, the heritable diversity within and between individuals and populations, was not a strategic research priority for Mediterranean forests during the decade 2011–2020 [34]. Keywords linked to forest genetics research remained rather inconspicuous in the bibliometric analysis of Nardi et al. [35] and were embedded within the research domain of biodiversity and conservation. However, genetic diversity is now widely recognized not only as underpinning ecosystem functioning, but also as strategic for biodiversity conservation and sustainable management [36, 37, 38, 39, 40••], including in the Mediterranean [25, 41]. Despite its foundational role in adaptation and resilience of ecosystems, genetic diversity has not received full attention in biodiversity research, as well as in natural resource management and policy-making worldwide [42, 43]. The indicators developed to facilitate biodiversity conservation and management based on genetic diversity are rarely adopted and genetic diversity is too rarely considered as a valid criterion for planning and designating protected areas [44, 45, 46].
Yet, forest tree genetic research worldwide has made very significant progress during the decade 2011–2020. Among others, new computational and genotyping/sequencing methods have emerged, species identification and population profiling have increased many folds, focus on the role of small-scale genetic diversity patterns in species adaptation and demography has risen sharply, and the importance of demography and selection in local to global patterns has been better understood [47, 48, 49, 50••].
Recognizing that in the context of global change, tree genetic diversity in the Mediterranean region represents a crucial resource for future forest adaptation; we review and highlight the major forest genetics research achievements of the past decades in Mediterranean countries. For this, we conducted a bibliometric analysis of the scientific literature covering the 30-year period (1991–2020) which follows the landmark Strasbourg conference on the protection of forests in Europe. The 1990 Strasbourg conference marked the beginning of an ongoing pan-European political process aiming at forest sustainable management. Through its resolution 2 (Conservation of Forest Genetic Resources), it led to the creation of EUFORGEN, the European Forest Genetic Resources Programme which has since then considerably helped promote forest gene conservation actions in Europe. Using this bibliometric analysis and putting together the representative regionwide expertise of our co-authorship, we address specifically the following: (i) what were the main forest genetics research trends in countries of the Mediterranean basin, in comparison with global trends, during the past thirty years; (ii) what are the research perspectives in the Mediterranean basin for the decade 2021–2030 to support the conservation and sustainable use of forest genetic resources; and (iii) what can the contribution of Mediterranean forest genetics research be to the global forest genetics research effort in the decade 2021–2030.
Forest Genetics Research for Conservation and Sustainable Use in the Countries of the Mediterranean Basin During the Period 1991–2020
Methods Used for the Bibliometric Analysis
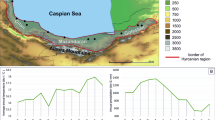
To quantitatively and qualitatively assess what were the main forest genetics research topics of the past decades in Mediterranean countries, and to identify trends, we performed a bibliometric analysis that was complemented by a qualitative analysis, based on our own expert knowledge, considering that bibliometric analyses do not capture a significant amount of knowledge produced in non-referenced scientific journals in national languages. Here, we define as Mediterranean countries those with at least part of their territory with a Mediterranean bioclimate [16••, 51] or those that belong to the FAO Committee on Mediterranean Forestry Questions – Silva Mediterranea (Fig. 1).

Countries investigated for their scientific production in the field of forest genetics during the period 1991–2020. The total number of scientific publications per country appears as a color scale using the specific Mediterranean query (QUERY2, see methods in Supplementary File S1)
Relevant scientific publications (not including grey literature) on forest tree genetics published during the period 1991–2020 were retrieved from the online Scopus database (https://www.scopus.com), October 11, 2021 [52].
The scientific literature was queried using the Scopus’ advanced search tool to find specific terms in an article’s title, abstract, and authors’ keywords [53]. The terms we used in the search queries were selected from a subset of 100 scientific publications, individually selected for their relevance to the field of forest genetics in the Southern European, west Asian, and north African part of the Mediterranean biome (i.e., the Mediterranean region [51]). We verified our selected keywords by running a preliminary query. We then enriched our list of keywords based on the additional terms that emerged from the preliminary query. We then further refined the list by excluding terms associated to other disciplines (medicine, zoology, etc.). The criteria followed for the selection of terms and the details of each query can be viewed in Supplementary File S1. We expect no significant bias from our search methodology (Supplementary File S1).
After obtaining a definitive list of search terms, we first carried out a general search of articles in order to run a comparative analysis between Mediterranean (QUERY1-1) and global (QUERY1-2) forest genetics research trends. We then performed a more refined search (QUERY2) of the publications of Mediterranean countries by incorporating into the query, country names (those of Fig. 1), and the tree genera of the Mediterranean region. Finally, we built QUERY3 to compare publication outputs in the field of forest genetics (QUERY2) with that of the field of forest ecology in Mediterranean countries. The details of the queries can be found in Supplementary File S1. The details of the filtering and validation processes can be found in Supplementary File S2. The details of the bibliometric analysis methods are in Supplementary File S3 while the thesaurus we created to curate our authors’ keyword database is in Supplementary File S4.
To assess the evolution of the research field over time in Mediterranean countries, the dataset of QUERY2 was split into the three sub-periods: 1991–2000; 2001–2010; 2011–2020, the latter one corresponding to the decade that the Mediterranean Forest Research Agenda (MFRA) of the European Forest Institute (EFI) addresses [34].
Trends in Publication Outputs
We first compared the total publication output on forest genetics of Mediterranean countries (QUERY1-1, 353 documents) with that of all countries combined (QUERY1-2, 12756 documents). Overall, the Mediterranean forest genetics research represents only 2.8% of the global output of publications on forest genetics. Figure 2 shows that during the first decade, Mediterranean countries accounted for 1.1% of the worldwide publication production in forest genetics (16 out of 1430). This fraction became 3.4% (119 out of 3507) in the second decade and decreased slightly to 2.8% in the last decade (218 out of 7819). Trends in publication rates increased at the same pace overall between 1991 and 2020 worldwide and in Mediterranean countries (Fig. 2). Mediterranean publications associated with the second decade, more specifically from 2003 to 2011 (Fig. 2), experienced an increased pace. It could be due to the discovery of new molecular biology technologies worldwide during the first decade of our analysis and implemented into Mediterranean research with a slight delay during the second decade, such as the very popular and efficient microsatellite markers (e.g., simple sequence repeat (SSR) [54]).

Trends in counts of published articles from 1991 to 2020 showing the increasing interest of the scientific community in forest genetics and comparing outputs from Mediterranean countries (in blue) and the World (in orange). Values on the Y axis represent, in percentage, the contribution of each year to the total publication output. Light colored lines are annual values while bold lines are moving average values calculated over the preceding 5 years. The box at the top left corner of the figure compares the absolute publication output in forest genetics per decade of Mediterranean countries with that of all countries in the world
The specific query for forest genetics research in Mediterranean countries (QUERY2) retrieved 1548 validated publications (2109 before filtering and validation). This scientific production is very modest compared to the other non-genetic domains (QUERY3, 40399 publications), representing only 3.8 to 5.2% of the total forest research output of Mediterranean countries.
The share of publications from Asia and Africa to the Mediterranean total increased during the 30-year period analyzed (Fig. 3). The increasing trend for west Asia and north Africa compared to Europe over the three decades could be due to an increase in the description of genetic resources of regional or local importance for forestry, food security, and health (e.g., [55, 56, 57]). During the last decade of 2011–2020, the number of publications for Mediterranean Asia, Africa, and Europe represented respectively the 21.6%, 7.9%, and 70.4% of the total amount of published articles (boxed graph in Fig. 3).

Trend of annual publications during the period 1991–2020 for each of the three continents of the Mediterranean region (Asia in blue, Africa in orange, and Europe in gray). Each annual value is calculated as a moving average over the past 5 years. Values on the Y axis represent, in percentage, the contribution of each year to the total publication output in each region. The smaller boxed graph shows the proportion of contribution per continent to the total of the Mediterranean publications in the three decades: 1991–2000; 2001–2010; 2011–2020
Major Scientific Topics of Interest in the Field of Mediterranean Forest Genetics
To assess the temporal trajectory of selected topics within the broader categories of scientific literature on forest genetics in Mediterranean countries, we first built a network map of keyword co-occurrences over the entire 30-year period (Fig. 4, see methods in Supplementary File S1) using QUERY2 (1548 publications). The minimum threshold defined for any given keyword to be included was its occurrence in at least 40 publications. Thus, 30 out of the 2945 keywords in the thesaurus were used to build a co-occurrence network (Fig. 4). These terms represent the most important topics addressed by the forest tree genetics community in Mediterranean countries. A similar number of keywords are often used in bibliometric analyses as it improves map visualization without losing significant information [58, 59]. Supplementary Table S1 reports the statistics associated to each keyword and cluster.

Co-occurrence network of the 30 most frequent authors’ keywords for the 30-year period (1991–2020). The distance between items demonstrates relative strength and topic similarity. Keywords belonging to the same cluster appear with the same color. The font and circle size of each keyword is proportional to the number of documents in which a keyword occurs; lines indicate co-occurrence links between terms, while line width is indicative of the link strength between two terms (number of publications in which two keywords occur together). The pie-chart at the bottom right of the figure shows the percentage contribution of each field to the total publications
Figure 4 indicates that forest genetics research in Mediterranean countries is organized into three different scientific domains of unequal total publication outputs: the red cluster 1 “Population diversity and Differentiation” with over 62% of the total number of publications, the green cluster 2 “Environmental conditions, growth and stress response” with almost 23%, and the blue cluster 3 “Phylogeography” with almost 15%.
Genetic diversity within population and differentiation among populations represent thus the major field of forest genetics research for the Mediterranean countries. Within this red cluster 1, the most frequent keywords were “Within-pop diversity,” “Differentiation,” “SSR,” and “Conservation” (Supplementary Table S1). Specifically, “Within-pop diversity” occupies a central position in the network (Fig. 4) suggesting that it frequently co-occurs with terms that belong to the other two clusters (highest total link strength of 852, Supplementary Table S1). On the contrary, the blue cluster 3, which refers to studies on biogeography and phylogeny, counts only 4 specific keywords out of the 30 selected. However, they score the highest citation rate with an average value of 55.13, suggesting that highly appreciated articles were published within this field and that they are foundational for the research community.
The green cluster 2 addresses topics associated with environmental conditions and adaptation; recurring terms are “Drought stress,” “Climate change,” “Growth,” and “Biodiversity.” Its lowest value of link strength (96.08) suggests that it is not as well connected as the other two clusters, representing a somewhat isolated topic of forest genetics research. This could be due to the fact that is the most recently investigated field of research (average of publication year is mid-2011, as opposed to early 2010 for the other two clusters, Supplementary Table S1). The keywords belonging to the green cluster 2, such as “climate change” and “drought stress,” have the most recent average year of publication (respectively, “2013.54” and “2013.07,” Supplementary Table S1).
In the following sub-sections, we analyze how topics deemed of high importance by our co-authorship are taken-up in publications and link to the main clusters and trends of the bibliometric analysis.
Exploring Understudied Genetic Resources: Genetic Diversity, Differentiation, and Marginal Populations
Throughout the period examined in our literature search, studies aimed at describing genetic diversity were initiated, updated, and completed in many ecologically and economically important Mediterranean and climatically sub-Mediterranean forest trees species, either widespread or of local interest, in some cases for the first time (e.g., [60, 61]). The three most studied species were also among the most economically important species in Southern Europe: Pinus pinaster, Quercus suber, and Pinus sylvestris (Fig. 5). They were targeted by 502 publications overall, corresponding to almost 33% of all Mediterranean research outputs found (1548). Studies were based on biochemical and modern molecular techniques, but also on more traditional morphometric research approaches mainly in common gardens. They included the study of growth and adaptation-related traits, in relation to climate change (Supplementary Table S1, Fig. 4). For the 10 most studied tree species presented in Fig. 5, the genetics literature (QUERY2) represented less than 8% of the total (QUERY3) on average, with a low 3% for Quercus ilex and a high 20% for Populus nigra (data not shown).

Changes in cumulative annual publications for the most studied forest tree species. The total in 2020 is the number of publications retrieved for a species during the period 1991–2020. The method for constructing the graph is explained in Supplementary File S4
Many marginal populations of widespread or endemic temperate species are found in the Mediterranean region. Marginal populations are characterized by habitats that are at the edges of species ranges, where conditions may be less suitable for survival and reproduction, and where specific adaptations presumably occur [62]. Thus, the conservation and sustainable use of these populations pose challenges that require the support of research. COST Action FP1202 (2012–2016) brought together research groups from Europe and Mediterranean countries interested in characterizing and exploring the genetic composition of these populations. It concluded that conservation policies should take marginal and peripheral populations into account, for their conservation and sustainable use. The adaptive potential of the genetic resources found in these populations was considered of relevance and importance for restoration projects and breeding programs [63].
Our bibliometric analysis shows that this topic has yet to gain importance. Although its average citation is within the range of citations of most keywords (Supplementary Table S1), the keyword “Marginal population” appears only in 13 articles with an average publication year centered on mid-2010 (Fig. 6). The keyword “Marginal population” is mostly connected with the red cluster 1 “Population diversity and differentiation” (box of Fig. 6, Fig. 4, Supplementary Table S1), but also to the blue cluster 3, “Phylogeography” (box of Fig. 6, Fig. 4). Seven out of the 13 publications connected to the “Marginal population” keyword concerned non-Mediterranean species, indicating an interest for populations located at the rear-edge (sensu [17]) of European distributions, as much as for marginality within the Mediterranean region itself (e.g., [64, 65, 66]). Finally, and interestingly, there was no marked connection with stress or climate related keywords (Fig. 6).

Co-occurrence network of the 30 most frequent authors’ keywords for the 30-year time frame (1991–2020) connected to the keyword “marginal populations,” with a time-trend perspective. Circle and link colors indicate the average publication year of the articles in which the relative keyword occurs. The font and circle size of each keyword is proportional to the number of documents in which a keyword occurs; lines indicate co-occurrence links between terms, while line width is indicative of the link strength between two terms (number of publications in which two keywords occur together). The table on the top-right of the figure reports statistics of the keywords to which the term “Marginal populations” is most related (see Supplementary Table S1 for a description of the headers)
The exploration of FGR also benefited from technological advances (see keywords in Fig. 4 and their date of average occurrence in Figs. 6 and 8). Moving from isozymes to highly polymorphic molecular markers such as SSR was widespread over the period examined, with research groups sharing the technical load of developing species-specific markers. Progress was also made in the field of genomics, benefiting from a worldwide technological trend. Yet, very few tree species genomes are fully described and sequenced, two of which are of Mediterranean species with agronomic importance: Prunus avium and Olea europaea [21, 67, 68, 69]. However, new genotyping/sequencing methods (exome capture, SPET technology, RADseq, etc.) were used in many Mediterranean species to characterize within and among population diversity and attempt at disentangling the effects of selection and drift on species and population evolution through time and space (e.g., [70, 71]).
Although not specific to the Mediterranean region, genetic monitoring has emerged as a key research subject, for example, to safeguard a declining biodiversity in EU countries (e.g., EU projects LifeGenMon and GenTree). Its conceptualization is based on a genecological framework and relies on the characterization of a minimum number of indicators and verifiers [72, 73]. Despite recent progress [74], many challenges remain, mainly in its implementation and acceptance by forest management and other stakeholders [75].
Genetic Improvement and Adaptation to Climate Change
The green cluster 2 (Fig. 4) is the most recently appeared cluster (mid-2011, Supplementary Table S1). It shows a clear connection between trait description, breeding, and adaptation (Supplementary Table S1). The keyword “Breeding” occurs 58 times and it is linked with 26 other terms (see Table of Fig. 7 for keywords with the strongest relationships). This connection varies in time, with breeding being more connected to population genetic descriptors during the decade 2011–2020, or disappearing entirely from the network during the decade 2001–2010 (average date for breeding is 2011.59). A main trend of change in genetic research has been from looking for promising genotypes for plantations to characterizing forest reproductive material (FRM) for the adaptation of forests to climate change and the mitigation of risks (Fig. 7). Thus, the link between breeding and drought resistance appears relatively late in our analysis (Fig. 7), indicating an increasing recognition that drought resistance is a key trait for the production of new forest reproductive material adapted to climate change.

Co-occurrence network of the 30 most frequent author keywords for the 30-year time frame (1991–2020) connected to the keyword “breeding,” with a time-trend perspective. Circle and link colors indicate the average publication year of the articles in which the relative keyword occurs. The font and circle size of each keyword is proportional to the number of documents in which a keyword occurs; lines indicate co-occurrence links between terms, while line width is indicative of the link strength between two terms (number of publications in which two keywords occur together). The table on the top-right of the figure reports statistics of the keywords to which the term “Breeding” is most related (see Supplementary Table S1 for a description of the headers)
Common gardens, where the phenotypic traits of FRM of different genetic origins are compared in a common environment, have been a major tool for testing adaptation to varying and changing environments, enabling breeding [76•]. Research breakthroughs during the last decade of the period analyzed made possible the revision of recommendations for use of FRM in some countries, so as to include climate change considerations, i.e., mostly where to source FRM that might best adapt to a changed mid-twenty-first century climate (for France, see https://agriculture.gouv.fr/graines-et-plants-forestiers-conseils-dutilisation-des-provenances-et-varietes-forestieres).
Most Mediterranean countries rely on natural regeneration and un-improved seed from identified seed stands for planting. However, breeding using multiple trait selection schemes and multiple environment testing is increasingly seen as a way to cope with climate change and to accelerate/anticipate adaptation measures (Fig. 7). Pinus pinaster remains the only example of a species with a long-developed breeding program (France, and more recently Italy, Morocco, Portugal, and Spain) backed by a strong stakeholder community [77]. Other breeding programs exist also for pines, such as, for example, for Pinus brutia (Turkey), Pinus halepensis (Greece, Morocco, Spain, and Turkey), Pinus pinea (Italy, Morocco, Portugal, Spain, Tunisia, and Turkey), P. nigra (France, Greece, Spain, and Turkey), and P. sylvestris (France, Greece, Spain, and Turkey). However, most of these breeding programs do not consider drought or pest resistance as their main target [78].
In low-input breeding programs, traditional and expensive long-term recurrent mating among elite lines is replaced by rapid selection of wild material using provenance tests and short-term surveys and/or easily acquired genomic data for phenotypic prediction or fingerprinting [42, 47, 79, 80]. Interest for low-input breeding is growing for numerous disseminated broadleaves such as Castanea sativa, Prunus avium, and Juglans regia, or conifers such as Pinus pinea [78]. Low-input breeding can be taken up by NGOs and can bring value to clonal archives. In France for example, seed orchards based on high diversity for increased resilience, rather than improved traits for production, built directly from provenance, progeny, or clonal tests, are emerging (for species such as Cedrus atlantica, Abies cephalonica, Pinus nigra subsp. salzmanni).
Phylogeography, Phylogeny, and Pleistocene History
Research on the evolutionary and demographic history of tree species present in the Mediterranean Basin, their Holocene colonization routes from refugia, and the identification of localized refugia in the Mediterranean refugial region (the “refugia-within-refugia” concept [81]), is the third major cluster of scientific interest for forest genetics in the Mediterranean (Fig. 4, blue cluster 3). This cluster developed as early as the red cluster 1 (early 2010, average date “2010.10,” Supplementary Table S1) and benefited strongly from the development of low effective population size and uniparentally inherited molecular markers [82], the formulation of the coalescent theory [83], and the development of computational tools (e.g., ABC simulations [84]). It is the most highly cited cluster of the three, probably because patterns described in Mediterranean tree species are applicable to other taxa and because the Mediterranean peninsulas were the glacial refugia of most European tree species [85].
Phylogeny also belongs to the blue cluster 3, probably because classical timed molecular phylogenies, based usually on low copy nuclear genes and/or mainly plastid DNA markers, have traditionally been used for reconstructing biogeographic patterns within geological time-frames [86, 87, 88]. During the period, and although paleoecology does not appear as a major keyword in Fig. 4, fossil pollen and/or macrofossil data were often coupled with phylogeographic surveys to investigate spatial distribution of extant genealogically linked lineages [89, 90, 91, 92, 93•] and ancient DNA was used to further reconstruct detailed evolutionary and demographic history of species [94, 95]. Also, phylogenetic approaches have been used in a spatial context to identify regions of high phylogenetic endemism [14•].
Interest in the investigation of phylogeographic patterns in Mediterranean tree species has slowed down over the 30-year period examined (Fig. 3). Yet, although refugial areas, often located in the Mediterranean [15, 96], and main Holocene colonization routes are known for European and sometimes for Mediterranean African and Asian species (e.g., [92, 97]), phylogeographic patterns are often not available at time scales long enough to identify significant events older than the Last Glacial Maximum [18••]. Neither are they available at spatial scales fine enough to help identify refugia and to assist with the design of conservation networks [89, 98]. They are also far from being available for all Mediterranean species [99].
Implications for Risk Mitigation, Conservation, and Sustainable Management
Forest fires constitute one of the main drivers of tree mortality and forest cover loss in Mediterranean countries [100]. The long-term impact of forest fires can explain speciation in Mediterranean trees [101] and the low genetic diversity found in thermophilic, obligatory seeding species, such as pines [23]. Also, a clear genetic signature of natural selection for serotiny, a major fire-response trait, has been demonstrated [102]. Yet, research on this topic has been limited and fire does not appear as a major keyword of the forest genetics literature (Fig. 4, Supplementary Table S1).
The keyword “Fire” is mostly connected with keywords of the green cluster 2. Its mean publication period is around late 2009 (Fig. 8), indicating a comparatively early, albeit restricted, interest in forest genetics. The keyword “Fire” occurred in only 10 articles and is linked to 29 other keywords; however, the strength of these connections is quite low. Figure 8 and its related table indicate a shift of research focus from selection and measurement of diversity to drought and climate change, potentially because of an increased interest in the correlation between drought and fire in a climate change context. Although weak, a link exists with the blue cluster 2 (see phylogeny in Fig. 8), indicating an interest for studying the role of fire and fire-response traits in evolutionary patterns of Mediterranean trees (e.g., [101]).

Co-occurrence network of the 30 most frequent author keywords for the 30-year time frame (1991–2020) connected to the keyword “fire,” with a time-trend perspective. Circle and link colors indicate the average publication year of the articles in which the relative keyword occurs. The font and circle size of each keyword is proportional to the number of documents in which a keyword occurs; lines indicate co-occurrence links between terms, while line width is indicative of the link strength between two terms (number of publications in which two keywords occur together). The table on the top-right of the figure reports statistics of the keywords to which the term “Fire” is most related (see Supplementary Table S1 for a description of the headers)
Under strong threats, ex situ conservation in the form of stands, conservation seed orchards, seed banks, or planted archives becomes a priority [103]. Yet, research on how to design core collections and manage long-term seed viability does not appear as a major keyword (Supplementary Table S1). Most countries manage sizeable networks of common gardens, dating from the 1960s to 1980s that were originally designed for breeding rather than conservation but can be considered as ex situ resources. There is scarce research on seed orchards that both conserve genetic resources ex situ and produce seeds for restoration projects or commercial plantations [104]. Finally, raised awareness of the importance of native genetic resources, possibly spurred by the adoption of the 2011 United Nations Nagoya protocol on Access and Benefit Sharing of genetic resources, has increased research on describing, mapping, conserving, and using native genetic resources [55, 56, 57, 60, 61, 63, 74], including for urban planning [105, 106, 107, 108].
Overall, “Within population diversity” and “Differentiation” (of the red cluster 1 “Population diversity and Differentiation,” Table 1) were the most frequently used keywords, keeping their respective first and third place across all three decades (Table 1). They were the corner stone of forest tree genetics research in the Mediterranean area, demonstrating an interest for both population level and landscape or range-wide genetic patterns and processes. Other keywords increased in frequency with time. The advancement of modern technologies and analytical potential can explain the exponential increase of the “SSR” term from 3% during the 1990–2000 decade to over 19% during the 2011–2020 decade (from 21st to second position, Table 1). During the last decade, keywords such as “Climate Change” and “Drought stress” (of the green cluster 2 “Environmental conditions, growth and stress response,” Table 1) also significantly increased their frequency (from 0.7% each during the 1990–2000 decade to 4.0% and 6.0% during the 2011–2020 decade, respectively), changing their ranking from 30th (both) to 10th and 6th, respectively (Table 1). Conversely, keywords related to biochemical techniques (isozymes), techniques proven unreliable (RAPD), or with limited genetic control (morphological traits) significantly decreased in frequency and ranking with time (Table 1).
Research Perspectives for the Decade 2021–2030 to Support the Conservation and Sustainable Use of Forest Genetic Resources
There are multiple threats affecting Mediterranean forest genetic resources. These threats are likely to continue increasing and intensifying. They include the following: climate change, leading to increased recurrent drought episodes, water vapor deficit, and heat waves; urban development, agriculture, and other land use changes associated with human demographic increase and resource mining, increased frequency and severity of disturbances, such as forest fragmentation, forest fires, and outbreak of pests and diseases [28]. During the past decades, forest genetics research has closely reflected these growing societal and environmental challenges (Supplementary Table S1). We now use the dynamics of past research interests evidenced by our bibliometric analysis as well as our own expectations and expertise as a basis for proposing research perspectives in the field of forest genetics for the upcoming decade.
Continue Exploring, Describing, and Characterizing Genetic Diversity of Poorly Known Species and Populations
Efforts at exploring, describing, and characterizing genetic diversity have certainly been carried out during the last decades, assessing genetic diversity within, and differentiation among populations, mostly for biogeographic and conservation purposes (Table 1, Fig. 4). Efforts have intensified in the Eastern and Southern Mediterranean during the most recent decade (Fig. 3). Yet, the hyper diverse tree flora of the Mediterranean basin [16••], which includes non-timber tree species of known or unknown economic value, is far from being fully explored, particularly in the Eastern and Southern Mediterranean. Thus, research should continue investigating fine scale species distribution and focus on their mapping (including threatened populations), taxonomic delineation (including genome-wide DNA barcode approaches), community composition, environmental variability, and genomic make-up [109]. The genetic diversity of under-used species and ecologically marginal and geographically peripheral populations, particularly, deserve further description of their genetic diversity, adaptive capacity, and phenotypic plasticity, thus bridging research interests from all three clusters (Fig. 4). Although proof is accumulating that they may contain rare, unique, and original adaptive genetic diversity [62] and that they may act as warning signals for emerging threats, their contribution to the overall diversity and resilience of species is still unclear.
Several benefits can be derived from the adoption of FGR management practices based on thorough genetic diversity descriptions. Better descriptions will improve science-based decision-making, for example, for determining areas where protection efforts (i.e., diversity hotspots) and restoration efforts (i.e., diversity cold spots) should increase [14•, 110]. Descriptions of multi-proxy patterns of diversity (taxonomic, functional, phylogenetic, genetic, e.g., [111]) will improve and optimize gene conservation networks [112••] and help selection of new material for breeding (“climate-smart” material), for example, in areas of high proxy congruence, in addition to guiding the sampling protocols for ex situ conservation in national gene banks. Better descriptions will also enable forest nurseries to diversify the FRM available for afforestation and restoration. Increasing the mapping of taxonomic and genetic diversity will also improve DNA tracking systems for testing the geographic origin and composition of FRM in nurseries and plantations, in the wood trading industry, for certification, and for detecting undesired exotic material in protected habitats. It will thus contribute to bridging the still wide knowledge transfer gap between forest genetics research, policymaking, and management [113].
Genomics of climate change traits should be the new direction, focusing on genes underlying traits such as drought, fire, insect and pest resistance and tolerance, increased phenotypic plasticity, and diversified microbiota—to improve our understanding of the genotype-phenotype-climate relationships and species adaptive space under changing environmental conditions (thus connecting more strongly the three research domains of Fig. 4). It is thus likely that, with the advent of the genome wide association studies (GWAS), interest in morphological traits will re-gain momentum (Table 1). What are the thresholds of collapse and extirpation remains a question of critical importance. Thus, better and faster phenotyping of traits meaningful for fitness and adaptation under rapidly changing conditions is the technological challenge to resolve, including the improvement of genomic predictions for breeding and conservation. In addition, the current genomic structure of natural populations holds precious information on the influence of past drivers of change. To learn from the past, probable demographic scenarios and signatures of selection should be tested in as many species as possible, regionally or range wide.
An enabling condition for the subsequent steps of testing (§ 3-2) and modeling (§ 3-3) is data availability. Yet, sharing data while protecting the intellectual property rights and sharing of benefits of data to resource owners is challenging. Sharing of archived data, broad collaboration, co-analysis of archived or neglected data, and open access databases are needed to draw new, solid, and generalizable conclusions, and so are research infrastructures connecting genetic diversity sites (GCUs, common gardens) with climate and environment monitoring devices. Transnational networking is needed more than ever (species habitats exist and move as climate changes across borders) at the scale of the Mediterranean basin, a non-politically integrated space where the support from regional institutions such as FAO Silva Mediterranea, EFIMED, and others is required. Conversely, data from management are strongly needed to upscale and broaden research findings [114]. Currently, such shared databases do not exist, and despite worthwhile efforts to archive data in open access repositories, forestry research and management are lagging behind. Data papers and creative commons licensing for old and new description data are a real opportunity for Mediterranean research and researchers that deserve recognition for their invaluable descriptive work (see for example the data papers published by the EU Horizon 2020 GenTree project [115, 116, 117]). Finally, there is a need for very precise metadata and to constantly improve data quality with clearly defined metadata standards with protocol descriptions and clearly defined data standards following minimum requirements (e.g., minimum sampling size, access to raw data for re-use).
Test and Experiment New Resources in New Environments Jointly Across Country Borders
Results from exploration, description, and characterization of genetic diversity are needed on their own. They are also needed for testing and experimenting new and little explored dimensions of forest tree species adaptation. Are today’s keystone species resilient? What is the pace and the spatial scale of local adaptation? How do demography and selection interact? To what extent do gene flow and hybridization which increase intraspecific genetic variation, facilitate, or prevent local adaptation? What role does the microbiota play in adaptation and to what extent is this microbiota affected by pollution (especially soil)? Such key questions require answers, not only because of their relevance for science, but also to improve species distribution models together with our understanding of resilience and its modification via silviculture [48, 49, 50, 118].
One of the methodological solutions for this is the planning and establishment of a new generation of common gardens with extended coverage of genetic diversity, habitats, and traits. High-resolution data such as those provided by satellite imagery are promising for accessing environmental information. Common gardens, although an old tool [119], remain particularly relevant as “space-for-time” experiments. While data from existing common gardens will continue to be of interest, particularly in the most challenging climates of Mediterranean Asia and Africa, we should acknowledge that they were designed and built at times of stable (or considered so) climate, mostly focusing on then promising genotypes for plantation forestry [42, 76•]. Thus, new common gardens are needed, testing new species, new lineages, and genotypes, in new and potentially harsher habitats, for new usage and traits of interest for the bioeconomy, not just for the Mediterranean region, but also in a broader temperate perspective as the Mediterranean climate is progressing to higher altitudes and latitudes under a climate-changed future [29, 120].
Testing risks in common gardens, including at early life stages in both nursery and natural conditions, is of key importance. Among recurring risks, and one that will likely increase, is drought stress, which went from being considered a topic of low concern to one of the top 10 topics of interest in the most recent decade (Table 1). Genomics of drought has moved from detecting single genes via random screening approaches [121] to not only identifying populations and lineages with potential adaptations, but also characterizing genes and gene networks involved in drought tolerance and resistance [76•, 122•]. Similarly, biotic interactions are likely to shift, increasing risks of insect and disease outbreaks. For example, bark beetles are generally considered as secondary pests that target stressed and weakened trees facing climate, soil, water, or pathogen stresses, but some species (primary pests) or some outbreaking populations of secondary pests can also attack and kill healthy trees [123]. This might lead to changing disturbance cycles, altered competition, and facilitation processes and possibly, to entirely new succession cycles and community composition.
New species and genotypes to test should be those likely to be “climate change winners” (including small and marginal populations as well as isolated trees, potentially adapted to extreme site condition, obtained from identified sources) and those likely to spread northward and to higher altitude. Also of importance are species for which the main interest may be other than wood production, such as those producing edible crops and health or green-chemistry-related compounds, or those with high ornamental value or high resistance to fire, erosion, or other biotic or abiotic risks. New genotypes for commercially important species should be selected from trees growing at ecological margins, both risking extirpation and being at the forefront of colonization. Habitats tested should also include ecological margins, while genetic material should be tested in replicated ways, including under controlled conditions in growth chambers and nurseries, to identify the factors driving early-stage genetic diversity patterns (demography and/or selection). The impact of nursery practices (culling, etc.) on the genetic variation of the produced FRM is also of concern and needs to be addressed.
Climate change “losers” should not be forgotten either. Whereas their habitat may shrink dramatically according to niche modeling, new areas will become favorable and genotypes and lineages that are likely to disappear under climate change in some areas may be the winners in other areas [124, 125•]. While genomic prediction and offset can be of help for such predictions [126], common garden experiments are needed as well for ground-proofing, both in habitats that are likely to disappear (for example in Mediterranean Asia and Africa), as well as in those that are likely to become favorable, so as to correctly and thoroughly test for general and local adaptations.
Finally, medium- to large-scale planting and restoration operations by the public and private sectors (forest companies), either conducted using local material or in the form of assisted migration plantations, using exotic or alternative provenance(s) in habitats of potential future suitability, can de facto be considered as valuable tests and demonstrators, to support upscaling common garden results, providing information on reproductive material selection (which origins succeed and which fail) and raising awareness of the importance of genetic diversity for adaptation. This exciting and original perspective of using everyday forestry practice as an experiment of adaptive potentials demands that passport (precise geographic origin of the material mostly) and performance data are safeguarded and shared. This is probably the most challenging element of this perspective.
Model, Simulate, and Build Scenarios to Inform Management and Policy for Mitigating and Adapting to Climate Change
Under the United Nations Framework Convention on Climate Change (UNFCCC) Paris Agreement of December 2015, forests worldwide are expected to provide both mitigation and adaptation solutions to anthropogenic disturbances. Yet, climate change is increasingly disrupting forests, sometimes severely in the form of massive die back, along with the benefits they provide to humans [127, 128]. Science is expected to provide solutions for their adaptive capacity, including via genetic adaptation, and to evaluate controversial management solutions that are starting to emerge such as assisted migration, species replacement and the use of non-native species, enrichment of natural forests via small scale plantation of expected climate change winners, composite provenancing, and targeted provenancing (see definitions in [129, 130]). All of these strategies to mitigate the expected effects of climate change contain some level of risk for the delivery of services, which science is asked and expected to evaluate [129]. These risks include biological invasions, disruption of local adaptation, and changes of dependent microbiota, maladaptation, and outbreeding depression.
In this context, forest tree species of the Mediterranean basin and their different gene pools are considered valuable genetic resources not just locally, but also for non-Mediterranean regions and countries, whose climate is predicted to evolve towards a Mediterranean-like climate by the end of the twenty-first century [131]. Thus, modeling the adaptive capacity of Mediterranean forest genetic resources has a far-reaching global interest. The modeling of current and expected species distribution ranges using ecological niche models that include processes (plasticity, phenology, dispersal, etc.) and increasingly complex and high-resolution environmental data [132, 133, 134] is one way of progressing in this domain.
Process-based biophysical models that take genetic diversity into consideration are also still needed to understand how genetic diversity, demography, and ecophysiological processes interact to mitigate collapse by water stress, pests and diseases, heat, frost, etc., all of which can occur over relatively short spatial and temporal scales [135]. This effort in the Mediterranean countries can be fueled mainly from monitoring of common garden data. Another area of development from such models is breeding. New generations of (low-) breeding programs incorporating diversity, resilience, and resistance to risks as an added (or alternate) goal to traditional biomass and wood production can and should emerge from an increased understanding of the phenotype–genotype–environment relationships and interactions. Finally, these models would also be very appropriate to test how forest management influences genetic diversity and vice versa, how genetic diversity benefits management for resilience, as experimenting in this field can be quite complicated (but see [136]).
Conclusion and Perspectives: Contribution of the Mediterranean Countries to the Global Forest Genetics Research Effort in the Decade 2021–2030
Forests are a defining element of both the natural landscape and the economy of Mediterranean countries, with genetic diversity a pillar of their biodiversity. Yet, research on forest genetics has remained modest in comparison to overall forest research in Mediterranean countries. During the next 10 years, the Mediterranean forest research community should have a prominent role in describing, testing, and modeling genetic diversity, understanding how severe threats due to climate change and other human activities can be mitigated, benefiting from high local and landscape heterogeneity (Fig. 9). One avenue for this could be to better cross-fertilize the currently compartmentalized three distinct research clusters that define the community of forest geneticists, based on our analysis, using common hypotheses and compatible methods, and linking results across clusters (Fig. 9). Its sharp local climate and ecological transitions make the Mediterranean region one of the most appropriate in the world to study microevolution (under different gene flow regimes), identify past and present drivers leading to the emergence of marginal populations, and evaluate the links between genotypes, phenotypes, and climatic drivers constraining tree growth, reproductive output and fitness, and, ultimately, adaptation [137, 138, 139, 140••].

Proposed research pathways in forest genetics for the management challenges of the 2021–2030 decade. The overarching goal of the research pathways is at the bottom of the figure while more specific research actions appear in higher levels of the figure. Blue boxes correspond to the “Continue exploring, describing and characterizing genetic diversity of poorly known species and populations” pathway (section 3-1), green boxes to the “Test and experiment new resources in new environments jointly across country borders” pathway (section 3-2), and yellow-brown boxes to the “Model, simulate and build scenarios to inform management and policy for mitigating and adapting to climate change” pathway (section 3-3)
There is a clear gap that needs filling between north, south, and east Mediterranean forest genetics knowledge. European forest tree species tend to be more fully described, tested, and modeled despite the importance of southern and eastern Mediterranean species in a region of the world with rapidly changing climate and strong pressures on biodiversity. The Mediterranean is not an integrated political arena and it does not benefit from a common framework that could shape agreements towards a common research goal. Integrating forest genetics research among EU and non-EU countries of the Mediterranean remains a challenge. Well-recognized institutions and programs, such as EUFORGEN (hosted by EFI), the PRIMA foundation, FAO Silva Mediterranea, the intergovernmental Union for the Mediterranean, and the European Union research framework programs, should take a leading role at fostering this research integration.
Mediterranean forest social-ecological systems have been shown to be particularly sensitive to forest policy management scenarios in their response to climate change [141, 142, 143, 144]. The lack of FGR-based management could be due both to missing knowledge and research, and to missing transfer of scientific knowledge to managers and policy makers [113]. The Mediterranean forest genetics research community thus has a leading role in meeting the needs of forest management based on the principles of multifunctional forest use, proposing sustainable management strategies for FGR in a highly heterogeneous landscape where forest ecosystems can be either resilient or declining. In doing so, the Mediterranean forest genetics research community will significantly contribute to many of the key commitments of EUFORGEN’s recently published European Forest Genetic Resources Strategy [145], including improving information on, conservation of, and sustainable use of forest genetic resources.
Finally, Mediterranean forests have been managed as multifunctional social-ecological systems for millennia, producing not just timber and fuel-wood but also many non-wood products and services, such as wild edible species important for food security [146, 147, 148]. As a multi-stakeholder social-ecosystem, forests are an ideal model for building participatory research approaches and study usage conflicts such as conservation with or without management and conservation vs exploitation [149], thus contributing to resolving the combined objectives of production, resilience, sustainability, and biodiversity protection. The Mediterranean Forest research community should increase efforts towards conducting cutting-edge multidisciplinary research and innovation in the area of sustainable management of FGR for traditional and new bioeconomy usage, under the pressure of climate change [150]. Multidisciplinary and transdisciplinary prospective work is needed, using IPCC scenarios as baselines, but moving beyond solutions limited to market economy paradigms which have up to now led to a rather unsustainable use of natural resources [151, 152]. The Mediterranean Forest research community can provide solutions to manage climate-induced risks and human pressure on Mediterranean forests. This needs to start with equitably sharing data, experience, and knowledge (Fig. 9).
Change history
28 September 2022
A Correction to this paper has been published: https://doi.org/10.1007/s40725-022-00173-y
Abbreviations
- AFLP:
-
Amplified fragment length polymorphism
- CBD:
-
Convention on Biological Diversity (https://www.cbd.int/)
- IUCN:
-
International Union for the Conservation of Nature (https://www.iucn.org/)
- EUFORGEN:
-
European Forest Genetic Resources Programme (http://www.euforgen.org/)
- EFI:
-
European Forest Institute (https://efi.int/)
- EFIMED:
-
EFI’s Mediterranean Facility (https://efi.int/efimed)
- FAO:
-
Food and Agriculture Organization of the United Nations (https://www.fao.org/)
- FOREST EUROPE:
-
The Ministerial Conference on the Protection of Forests in Europe (https://foresteurope.org/)
- FSC:
-
Forest Stewardship Council (https://fsc.org/en)
- FRM:
-
Forest reproductive material
- FGR:
-
Forest genetic resources
- GCU:
-
Gene conservation unit
- GR:
-
Genetic resources
- MFRA:
-
Mediterranean Forest Research Agenda of EFI-Med
- NGO:
-
Non-governmental organization
- PEFC:
-
Programme for the Endorsement of Forest Certification (https://www.pefc.org/)
- PRIMA Foundation:
-
Partnership for Research and Innovation in the Mediterranean Area (https://prima-med.org/)
- RAPD:
-
Random amplified polymorphic DNA
- SSR:
-
Simple sequence repeat
References
Papers of particular interest, published recently, have been highlighted as: • Of importance •• Of major importance
••Pörtner HO, Scholes RJ, Agard J, Archer E, Arneth A, Bai X, et al. Scientific outcome of the IPBES-IPCC co-sponsored workshop on biodiversity and climate change. Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES). https://zenodo.org/record/5101125, 2021. This joint IPBES-IPCC report indicates the urgency of taking mitigating and adaptation measures jointly for human societies and biodiversity.
••FAO, UNEP. The State of the World’s Forests 2020. Forests, biodiversity and people. Rome, 2021. This is an updated reference assessment of the state of the world’s forests, with key figures and trends. The report’s definition of forests which we adopted is land spanning more than 0.5 hectares with trees higher than 5 meters and a canopy cover of more than 10 percent, or trees able to reach these thresholds in situ, excluding land predominantly under agricultural or urban land use.
CBD, Secretariat of the Convention on Biological Diversity. Global Biodiversity Outlook 5, Montréal, 2020.
Turney C, Ausseil AG, Broadhurst L. Urgent need for an integrated policy framework for biodiversity loss and climate change. Nature Ecology & Evolution. 2020;4(8):996–6.
IPCC. Summary for policymakers. In: Masson-Delmotte V, Zhai P, Pirani A, Connors SL, Péan C, Berger S, et al., editors. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change: Cambridge University Press; 2021.
Mace GM, Barrett M, Burgess ND, Cornell SE, Freeman R, Grooten M, Purvis A. Aiming higher to bend the curve of biodiversity loss. Nature Sustainability. 2018;1(9):448–51.
Mazor T, Doropoulos C, Schwarzmueller F, Gladish DW, Kumaran N, Merkel K, et al. Global mismatch of policy and research on drivers of biodiversity loss. Nature Ecology & Evolution. 2018;2(7):1071–4.
Quézel P. Analysis of the flora of Mediterranean and Saharan Africa. Annals of the Missouri Botanical Garden. 1978;65:479–534.
Myers N, Mittermeier RA, Mittermeier CG, Da Fonseca GA, Kent J. Biodiversity hotspots for conservation priorities. Nature. 2000;403(6772):853–8.
Medail F, Quézel P. Hot-spots analysis for conservation of plant biodiversity in the Mediterranean Basin. Annals of the Missouri Botanical Garden. 1997;84:112–27.
Cañadas EM, Fenu G, Peñas J, Lorite J, Mattana E, Bacchetta G. Hotspots within hotspots: endemic plant richness, environmental drivers, and implications for conservation. Biological Conservation. 2014;170:282–91.
Cuttelod A, García N, Malak DA, Temple HJ, Katariya V. The Mediterranean: a biodiversity hotspot under threat. Wildlife in a Changing World–an analysis of the 2008 IUCN Red List of. Threatened Species. 2009;89(2019):9.
Thompson JD. Plant evolution in the Mediterranean. Oxford: Oxford University Press; 2005.
•Cheikh AM, Escudero M, Monnet AC, Arroyo J, Bacchetta G, Bagnoli F, et al. Spatial patterns of genus-level phylogenetic endemism in the tree flora of Mediterranean Europe. Diversity and Distributions. 2021;27(5):913–28. This paper builds on the idea of phylogenetic endemism to identify regions of high biodiversity (and thus conservation) value in the Mediterranean region.
Médail F, Diadema K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. Journal of Biogeography. 2009;36:1333–45.
••Médail F, Monnet AC, Pavon D, Nikolic T, Dimopoulos P, Bacchetta G, et al. What is a tree in the Mediterranean Basin hotspot? A critical analysis. Forest Ecosystems. 2019;6:17. The first and only checklist of tree species in the European part of the Mediterranean region.
Hampe A, Petit RJ. Conserving biodiversity under climate change: the rear edge matters. Ecology Letters. 2005;8(5):461–7.
••Magri D, Di Rita F, Aranbarri J, Fletcher W, González-Sampériz P. Quaternary disappearance of tree taxa from Southern Europe: timing and trends. Quaternary Science Reviews. 2017;163:23–55. A paleo-ecological analysis of the disappearance of taxa from local floras across southern Europe, proving that many disappearance events occurred earlier than the often-invoked Last Glacial Maximum.
Aravanopoulos FA. Do silviculture and forest management affect the genetic diversity and structure of long-impacted forest tree populations? Forests. 2018;9(355):1–14.
Cheikh AM, Escudero M, Ponger L, Monnet AC, Arroyo J, Nikolic T, et al. A comprehensive, genus-level time-calibrated phylogeny of the tree flora of Mediterranean Europe and an assessment of its vulnerability. Botany Letters. 2020;167:276–89.
Pinosio S, Marroni F, Zuccolo A, Vitulo N, Mariette S, Sonnante G, et al. A draft genome of sweet cherry (Prunus avium L.) reveals genome-wide and local effects of domestication. The Plant Journal. 2020;103:1420–32.
Fady B. Is there really more biodiversity in Mediterranean forest ecosystems? Taxon. 2005;54(4):905–10.
Fady B, Conord C. Macroecological patterns of species and genetic diversity in vascular plants of the Mediterranean Basin. Diversity and Distributions. 2010;16(1):53–64.
Rundel PW. A Neogene heritage: conifer distributions and endemism in Mediterranean-climate ecosystems. Frontiers in Ecology and Evolution. 2019;7:364.
Scarascia-Mugnozza G, Oswald H, Piussi P, Radoglou K. Forests of the Mediterranean region: gaps in knowledge and research needs. Forest Ecology and Management. 2000;132(1):97–109.
Gauquelin T, Michon G, et al., Joffre R, Duponnois R, Génin D, Fady B, et al. Mediterranean forests, land use and climate change: a social-ecological perspective. Regional Environmental Change. 2018;18:623–36.
Quézel P, Médail F. Ecologie et biogéographie des forêts du bassin méditerranéen. Paris, France: Elsevier; 2003.
FAO. State of Mediterranean Forests. Rome, Italy; 2018.
MedECC. Climate and environmental change in the Mediterranean Basin – current situation and risks for the future. In: Cramer W, Guiot J, Marini K, editors. First Mediterranean Assessment Report. Marseille, France: Union for the Mediterranean, Plan Bleu, UNEP/MAP; 2020.
Millar CI, Stephenson NL. Temperate forest health in an era of emerging megadisturbance. Science. 2015;349(6250):823–6.
Sala OE, Chapin FS, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, et al. Global biodiversity scenarios for the year 2100. Science. 2000;287(5459):1770–4.
Kovats RS, Valentini R, Bouwer LM, Georgopoulou E, Jacob D, Martin E, Rounsevell M, Europe SJF. In: Barros VR, Field CB, Dokken DJ, Mastrandrea MD, Mach KJ, Bilir TE, et al., editors. Climate change 2014: impacts, adaptation, and vulnerability. Part B: Regional aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press; 2014. p. 1267–326.
Concepción ED. Urban sprawl into Natura 2000 network over Europe. Conservation Biology. 2021;35:1063–72.
EFIMED. A Mediterranean Forest Research Agenda (MFRA) for the period 2010-2020. Barcelona, Spain: European Forest Institute; 2009.
Nardi P, Di Matteo G, Palahi M, Scarascia MG. Structure and evolution of Mediterranean forest research: a science mapping approach. PLoS ONE. 2016;11(5):e0155016.
Jump AS, Marchant R, Peñuelas J. Environmental change and the option value of genetic diversity. Trends in Plant Science. 2009;14(1):51–8.
Liang J, Crowther TW, Picard N, Wiser S, Zhou M, Alberti G, et al. Positive biodiversity-productivity relationship predominant in global forests. Science. 2016;354(6309):aaf8957.
Lefèvre F, Boivin T, Bontemps A, Courbet F, Davi H, Durand-Gillmann M, et al. Considering evolutionary processes in adaptive forestry. Annals of Forest Science. 2014;71:723–39.
Ducci F. Genetic resources and forestry in the Mediterranean region in relation to global change. Annals of Silvicultural Research. 2015;39(2):70–93.
••Hoban S, Bruford M, Jackson JDU, Lopes-Fernandes M, Heuertz M, Hohenlohe PA, et al. Genetic diversity targets and indicators in the CBD post-2020 Global Biodiversity Framework must be improved. Biological Conservation. 2020;248:108654. A much needed but debated proposal for including genetic diversity indicators in biodiversity assessment frameworks.
Bariteau M, Alptekin U, Aravanopoulos FA, Asmar F, Bentouati A, Benzyane M, et al. Les resources génétiques forestières dans le basin méditerranéen. Forêt Méditerranéenne. 2003;24(2):148–58.
Fady B, Cottrell J, Ackzell L, Alía R, Muys B, Prada A, González-Martínez SC. Forests and global change: what can genetics contribute to the major forest management and policy challenges of the twenty-first century? Regional Environmental Change. 2016;16(4):927–39.
Hoban S, Campbell CD, da Silva JM, Ekblom R, Funk WC, Garner BA, et al. Genetic diversity is considered important but interpreted narrowly in country reports to the Convention on Biological Diversity: Current actions and indicators are insufficient. Biological Conservation. 2021;261:109233.
Graudal L, Aravanopoulos F, Bennadji Z, Changtragoon S, Fady B, Kjær ED, et al. Global to local genetic diversity indicators of evolutionary potential in tree species within and outside forests. Forest Ecology & Management. 2014;333:35–51.
Lefèvre F, Alía R, Bakkebø FK, Graudal L, Oggioni SD, Rusanen M, Vendramin GG, Bozzano M. Dynamic conservation and utilization of forest tree genetic resources: indicators for in situ and ex situ genetic conservation and forest reproductive material. European Forest Genetic Resources Programme (EUFORGEN). Barcelona, Spain: European Forest Institute; 2020.
Fady B, Bozzano M. Effective population size does not make a practical indicator of genetic diversity in forest trees. Biological Conservation. 2021;253:108904.
Neale D, Kremer A. Forest tree genomics: growing resources and applications. Nature Reviews Genetics. 2011;2(2):111–22.
Plomion C, Bastien C, Bogeat-Triboulot MB, Bouffier L, Déjardin A, Duplessis S, et al. Forest tree genomics: 10 achievements from the past 10 years and future prospects. Annals of Forest Science. 2016;73:77–103.
Holliday J, Aitken S, Cooke JE, Fady B, González-Martínez SC, Heuertz M, et al. Advances in ecological genomics in forest trees and applications to genetic resources conservation and breeding. Molecular Ecology. 2017;26:706–17.
••Fady B, Aravanopoulos F, Benavides R, González-Martínez S, Grivet D, Lascoux M, et al. Genetics to the rescue: managing forests sustainably in a changing world. Tree Genetics & Genomes. 2020;16(6):1–11. The latest review highlighting genetic research topics of high importance for sustainable forest management.
Olson DM, Dinerstein E, Wikramanayake ED, Burgess ND, Powell GV, Underwood EC, et al. Terrestrial ecoregions of the world: a new map of life on Earth. A new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. BioScience. 2001;51(11):933–8.
Mongeon P, Paul-Hus A. The journal coverage of Web of Science and Scopus: a comparative analysis. Scientometrics. 2016;106:213–28.
Berkvens P. Scopus custom data documentation. Retrieved from https://p.widencdn.net/mrbekb/Scopus_Custom_Data_ Documentation_Version9, 2012.
Morgante M, Olivieri AM. PCR-amplified microsatellites as markers in plant genetics. The Plant Journal. 1993;3(1):175–82.
Shahghobadi H, Shabanian N, Rahmani MS, Khadivi A. Genetic characterization of Pistacia atlantica subsp. kurdica from northern Zagros forests in Iran. Trees. 2019;33(2):481–90.
Douaihy B, Vendramin GG, Boratyński A, Machon N, Bou Dagher-Kharrat M. High genetic diversity with moderate differentiation in Juniperus excelsa from Lebanon and the eastern Mediterranean region. AoB Plants. 2011:plr003.
Nasri N, Tlili N, Ammar KB, Khaldi A, Fady B, Triki S. High tocopherol and triacylglycerol contents in Pinus pinea L. seeds. International Journal of Food Sciences and Nutrition. 2009;60:161–9.
Aznar-Sánchez JA, Belmonte-Ureña LJ, López-Serrano MJ, Velasco-Muñoz JF. Forest ecosystem services: an analysis of worldwide research. Forests. 2018;9(8):453.
Ho YS, Mukul SA. Publication Performance and Trends in Mangrove Forests: A Bibliometric Analysis. Sustainability. 2021;13(22):12532.
Awad L, Fady B, Khater C, Roig A, Cheddadi R. Genetic structure and diversity of the endangered fir tree of Lebanon (Abies cilicica Carr.): implications for conservation. PLoS One. 2014;9(2):e90086.
Walas Ł, Sobierajska K, Ok T, Dönmez AA, Kanoğlu SS, Bou Dagher-Kharrat MB, et al. Past, present, and future geographic range of an oro-Mediterranean Tertiary relict: the Juniperus drupacea case study. Regional Environmental Change. 2019;19(5):1507–20.
Csilléry K, Buchmann N, Fady B. Adaptation to drought is coupled with slow growth, but independent from phenology in marginal silver fir (Abies alba Mill.) populations. Evolutionary Applications. 2020;13(9):2357–76.
Ducci F, Cutino I, Monteverdi MC, Picard N, Proietti R, et al. Marginal/peripheral populations of forest tree species and their conservation status: report for Mediterranean region. Annals of Silvicultural Research. 2017;41:31–40.
Moracho E, Moreno G, Jordano P, Hampe A. Unusually limited pollen dispersal and connectivity of pedunculate oak (Quercus robur) refugial populations at the species’ southern range margin. Molecular Ecology. 2016;25(14):3319–31.
Piotti A, Leonarduzzi C, Postolache D, Bagnoli F, Spanu I, Brousseau L, et al. Unexpected scenarios from Mediterranean refugial areas: disentangling complex demographic dynamics along the Apennine distribution of silver fir. Journal of Biogeography. 2017;44(7):1547–58.
de Dato GD, Teani A, Mattioni C, Aravanopoulos F, Avramidou EV, Stojnic S, et al. Genetic analysis by nuSSR markers of silver Birch (Betula pendula Roth) populations in their Southern European distribution range. Frontiers in Plant Science. 2020;11:310.
Cruz F, Julca I, Gómez-Garrido J, Loska D, Marcet-Houben M, Cano E, et al. Genome sequence of the olive tree, Olea europaea. GigaScience. 2016;5(1):s13742–016.
Shirasawa K, Isuzugawa K, Ikenaga M, Saito Y, Yamamoto T, Hirakawa H, Isobe S. The genome sequence of sweet cherry (Prunus avium) for use in genomics-assisted breeding. DNA Research. 2017;24(5):499–508.
Wang J, Liu W, Zhu D, Zhou X, Hong P, Zhao H, et al. A de novo assembly of the sweet cherry (Prunus avium cv. Tieton) genome using linked-read sequencing technology. PeerJ. 2020;8:e9114.
Karam MJ, Lefèvre F, Bou Dagher-Kharrat M, Pinosio S, Vendramin GG. Genomic exploration and molecular marker development in a large and complex conifer genome using RADseq and mRNAseq. Molecular Ecology Resources. 2015;15(3):601–12. https://doi.org/10.1111/1755-0998.12329.
Houminer N, Doron-Faigenboim A, Shklar G, De La Torre AD, Neale D, Korol L. Transcriptome-based single-nucleotide polymorphism markers between Pinus brutia and Pinus halepensis and the analysis of their hybrids. Tree Genetics & Genomes. 2021;17:14.
Aravanopoulos FA. Genetic monitoring in natural perennial plant populations. Botany. 2011;89:75–81.
Aravanopoulos F.A., Tollefsrud M.M., Graudal L., Koskela J., Kätzel R., Soto A., et al. Development of genetic monitoring methods for genetic conservation units of forest trees in Europe. European Forest Genetic Resources Programme (EUFORGEN), Bioversity International, Rome, Italy, 2015, 62 pp.
Bajc M, Aravanopoulos F, Westergren M, Fussi B, Kavaliauskas D, Alizoti P, et al. Manual for forest genetic monitoring. Ljubljana, 332 pp.: Silva Slovenica Publishing Centre; 2020. https://doi.org/10.20315/SFS.167.
GenRes Bridge Project Consortium, ECPGR, ERFP and EUFORGEN. Genetic Resources Strategy for Europe. European Forest Institute, 2021, 64 p.
•Ramírez-Valiente J.A., Santos del Blanco L., Alía R., Robledo-Arnuncio J. J., Climent J. Adaptation of Mediterranean forest species to climate: lessons from common garden experiments. Journal of Ecology. 2021;110(5):1022–1042. A review and analysis of common garden data showing that no negative association between growth- and persistence-related traits at the intraspecific level, but marked adaptation to temperature and precipitation regimes.
Bouffier L, Raffin A, Alia R. Maritime pine – Pinus pinaster Ait. In: Mullin TJ, Lee SJ, editors. Best practice for tree breeding in Europe. Gävle: Skogforsk; 2013. p. 65–76.
Climent J, Alizoti P, Rodriguez-Quilón I, Kurt Y, Ducci F, Fady B, Alía R. Conservation and breeding of Mediterranean pines. In: Ne’eman G, Osem Y, editors. Pines and their mixed forest ecosystems in the Mediterranean Basin. Managing Forest Ecosystems, vol. vol 38. Cham: Springer; 2021.
Wang XR, Torimaru T, Lindgren D, Fries A. Marker-based parentage analysis facilitates low input ‘breeding without breeding’ strategies for forest trees. Tree Genetics & Genomes. 2010;6(2):227–35.
El-Kassaby Y, Lstibůrek M. Breeding without breeding. Genetics Research. 2009;91:111–20.
Gómez A, Lunt DH. Refugia within refugia: patterns of phylogeographic concordance in the Iberian Peninsula. In: Weiss S, Ferrand N, editors. Phylogeography of southern European refugia. Berlin: Springer; 2007. p. 155–88.
Avise JC. Phylogeography: the history and formation of species. Cambridge: Harvard University Press; 2000.
Kingman JFC. The coalescent. Stochastic Processes and their Applications. 1982;13(3):235–48.
Beaumont MA, Zhang W, Balding DJ. Approximate Bayesian computation in population genetics. Genetics. 2002;162:2025–35.
Petit RJ, Aguinagalde I, de Beaulieu JL, Bittkau C, Brewer S, Cheddadi R, et al. Glacial refugia: hotspots but not melting pots of genetic diversity. Science. 2003;300(5625):1563–5.
Qiao CY, Ran JH, Li Y, Wang XQ. Phylogeny and biogeography of Cedrus (Pinaceae) inferred from sequences of seven paternal chloroplast and maternal mitochondrial DNA regions. Annals of Botany. 2007;100(3):573–80.
Gömöry D, Paule L, Macejovsky V. Phylogeny of beech in western Eurasia as inferred by approximate Bayesian computation. Acta Societatis Botanicorum Poloniae. 2018;87(2):3582.
Sękiewicz K, Dering M, Romo A, Dagher-Kharrat MB, Boratyńska K, Ok T, Boratyński A. Phylogenetic and biogeographic insights into long-lived Mediterranean Cupressus taxa with a schizo-endemic distribution and Tertiary origin. Botanical Journal of the Linnean Society. 2018;188(2):190–212.
Petit RJ, Brewer S, Bordács S, Burg K, Cheddadi R, Coart E, et al. Identification of refugia and post-glacial colonisation routes of European white oaks based on chloroplast DNA and fossil pollen evidence. Forest ecology and Management. 2002;156(1-3):49–74.
Hampe A, Arroyo J, Jordano P, Petit RJ. Rangewide phylogeography of a bird-dispersed Eurasian shrub: contrasting Mediterranean and temperate glacial refugia. Molecular Ecology. 2003;12(12):3415–26.
Cheddadi R, Vendramin GG, Litt T, François L, Kageyama M, Lorentz S, et al. Imprints of glacial refugia in the modern genetic diversity of Pinus sylvestris. Global Ecology and Biogeography. 2006;15(3):271–82.
Cheddadi R, Fady B, François L, Hajar L, Suc JP, Huang K, et al. Putative glacial refugia of Cedrus atlantica deduced from Quaternary pollen records and modern genetic diversity. Journal of Biogeography. 2009;36(7):1361–71.
•Besnard G, Khadari B, Navascués M, Fernández-Mazuecos M, El Bakkali A, Arrigo N, et al. The complex history of the olive tree: from Late Quaternary diversification of Mediterranean lineages to primary domestication in the northern Levant. Proceedings of the Royal Society B: Biological Sciences. 2013;280(1756):20122833. A classic and well-designed study exemplifying the interplay between natural and domestication demographic events in shaping phylogeographic patterns in the Mediterranean.
Parducci L, Petit RJ. Ancient DNA–unlocking plants’ fossil secrets. New Phytologist. 2004;161(2):335–9.
Hu FS, Hampe A, Petit RJ. Paleoecology meets genetics: deciphering past vegetational dynamics. Frontiers in Ecology and the Environment. 2009;7:371–9.
Tzedakis PC, Emerson BC, Hewitt GM. Cryptic or mystic? Glacial tree refugia in northern Europe. Trends in Ecology and Evolution. 2013;28(12):696–704.
Mattioni C, Martin MA, Pollegioni P, Cherubini M, Villani F. Microsatellite markers reveal a strong geographical structure in European populations of Castanea sativa (Fagaceae): evidence for multiple glacial refugia. American Journal of Botany. 2013;100(5):951–61.
Dering M, Kosiński P, Wyka TP, Pers-Kamczyc E, Boratyński A, Boratyńska K, et al. Tertiary remnants and Holocene colonizers: genetic structure and phylogeography of Scots pine reveal higher genetic diversity in young boreal than in relict Mediterranean populations and a dual colonization of Fennoscandia. Diversity and Distributions. 2017;23:540–55.
Le VJ, Galliot N, Pironon S, Nieto FG, Suc JP, Lakhal-Mirleau F, et al. A strong east–west Mediterranean divergence supports a new phylogeographic history of the carob tree (Ceratonia siliqua, Leguminosae) and multiple domestications from native populations. Journal of Biogeography. 2020;47(2):460–71.
Klein T, Cahanovitc R, Sprintsin M, Herr N, Schiller G. A nation-wide analysis of tree mortality under climate change: forest loss and its causes in Israel 1948–2017. Forest Ecology and Management. 2019;432:840–9.
Verdú M, Pausas JG. Fire drives phylogenetic clustering in Mediterranean Basin woody plant communities. Journal of Ecology. 2007;95(6):1316–23.
Budde KB, Heuertz M, Hernández-Serrano A, Pausas JG, Vendramin GG, Verdú M, González-Martínez SC. In situ genetic association for serotiny, a fire-related trait, in Mediterranean maritime pine (Pinus pinaster). New Phytologist. 2014;201(1):230–41.
Skrøppa T. Ex situ conservation methods. In: Conservation and management of forest genetic resources in Europe. Arbora Publishers, Zvolen, 2005, p567-583.
Eliades NGH, Papageorgiou AC, Fady B, Gailing O, Leinemann L, Finkeldey R. An approach to genetic resources conservation of peripheral isolated plant populations: the case of an island narrow endemic species. Biodiversity and Conservation. 2019;28(11):3005–35.
Alvey AA. Promoting and preserving biodiversity in the urban forest. Urban Forestry & Urban Greening. 2006;5(4):195–201.
Vimal R, Geniaux G, Pluvinet P, Napoléone C, Lepart J. Detecting threatened biodiversity by urbanization at regional and local scales using an urban sprawl simulation approach: application on the French Mediterranean region. Landscape and Urban Planning. 2012;104(3-4):343–55.
Capotorti G, del Vico E, Lattanzi E, Tilia A, Celesti-Grapow L. Exploring biodiversity in a metropolitan area in the Mediterranean region: the urban and suburban flora of Rome (Italy). Plant Biosystems. 2013;147(1):174–85.
Ballesteros D, Meloni F, Bacchetta G, editors. Manual for the propagation of selected Mediterranean native plant species: Ecoplantmed, ENPI, CBC-MED; 2015.
Monnet AC, Cilleros K, Médail F, Albassatneh MC, Arroyo J, Bacchetta G, et al. WOODIV, a database of occurrences, functional traits, and phylogenetic data for all Euro-Mediterranean trees. Scientific Data. 2021;8:89.
Morelli TL, Barrows CW, Ramirez AR, Cartwright JM, Ackerly DD, Eaves TD, et al. Climate-change refugia: biodiversity in the slow lane. Frontiers in Ecology and the Environment. 2020;18(5):228–34.
Doxa A, Devictor V, Baumel A, Pavon D, Medail F, Leriche A. Beyond taxonomic diversity: Revealing spatial mismatches in phylogenetic and functional diversity facets in Mediterranean tree communities in southern France. Forest Ecology and Management. 2020;474:118318.
••Lefèvre F, Koskela J, Hubert J, Kraigher H, Longauer R, Olrik DC, et al. Dynamic conservation of forest genetic resources in 33 European countries. Conservation Biology. 2013;27(2):373–84. This paper details the bases of in situ gene conservation as it is practiced and implemented in Europe.
Vinceti B, Manica M, Lauridsen N, Verkerk PJ, Lindner M, Fady B. Managing forest genetic resources as a strategy to adapt forests to climate change: perceptions of European forest owners and managers. European Journal of Forest Research. 2020;139(6):1107–19.
Fady B, Geburek T, Scotti I. Science needs management data for a better prediction of climate change effects on socio-ecosystems. Annals of Forest Science. 2019;76(1):12.
Benavides R, Carvalho B, Bastias CC, López-Quiroga D, Mas A, Cavers S, et al. The GenTree Leaf Collection: inter- and intraspecific leaf variation in seven forest tree species in Europe. Global Ecology and Biogeography. 2021;30(3):590–7.
Martínez-Sancho E, Slámová L, Morganti S, Grefen C, Carvalho B, Dauphin B, et al. The GenTree Dendroecological Collection, tree-ring and wood density data from seven tree species across Europe. Scientific Data. 2020;7:1–7.
Opgenoorth L, Dauphin B, Benavides R, Heer K, Alizoti P, Martínez-Sancho E, et al. The GenTree Platform: growth traits and tree-level environmental data in 12 European forest tree species. GigaScience. 2021;10(3):giab010.
Rius M, Darling JA. How important is intraspecific genetic admixture to the success of colonising populations? Trends in Ecology & Evolution. 2014;29(4):233–42.
Langlet O. Two hundred years genecology. Taxon. 1971;20(5-6):653–721.
Fady B, Rihm G. Arboretums, common gardens and forest tree resilience. New Forests. 2022;53:603–606.
Jump AS, Hunt JM, Martinez-Izquierdo JA, Peñuelas J. Natural selection and climate change: temperature-linked spatial and temporal trends in gene frequency in Fagus sylvatica. Molecular Ecology. 2006;15(11):3469–80.
•Martínez-Sancho E, Rellstab C, Guillaume F, Bigler C, Fonti P, Wohlgemuth T, Vitasse Y. Post-glacial re-colonization and natural selection have shaped growth responses of silver fir across Europe. Science of the Total Environment. 2021;779:146393. This paper suggests that both post-glacial colonization and natural selection are the major drivers of within species variability in growth in a common European forest tree species (silver fir).
Raffa KF, Powell EN, Townsend PA. Temperature-driven range expansion of an irruptive insect heightened by weakly coevolved plant defenses. Proceedings of the National Academy of Sciences USA. 2013;110:2193–8.
Dyderski MK, Paź S, Frelich LE, Jagodziński AM. How much does climate change threaten European forest tree species distributions? Global Change Biology. 2018;24(3):1150–63.
•Benito GM, Alía R, Robson TM, Zavala MA. Intra-specific variability and plasticity influence potential tree species distributions under climate change. Global Ecology and Biogeography. 2011;20(5):766–78. One of the first papers showing that not including biological processes (here phenotypic plasticity) in niche modeling leads to strong habitat suitability biases.
Rellstab C, Dauphin B, Exposito-Alonso M. Prospects and limitations of genomic offset in conservation management. Evolutionary Applications. 2021;14(5):1202–12.
Lindner M, Maroschek M, Netherer S, Kremer A, Barbati A, Garcia-Gonzalo J, et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. Forest Ecology and Management. 2010;259(4):698–709.
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest ecology and management. 2010;259(4):660–84.
Prober SM, Byrne M, McLean EH, Steane DA, Potts BM, Vaillancourt RE, Stock WD. Climate-adjusted provenancing: a strategy for climate-resilient ecological restoration. Frontiers in Ecology and Evolution. 2015;3:65.
Notivol E, Santos-Del-blanco L, Chambel R, Climent J, Alía R. Seed sourcing strategies considering climate change forecasts: a practical test in Scots pine. Forests. 2020;11(11):1–17.
Giorgi F, Bi X, Pal J. Mean, interannual variability and trends in a regional climate change experiment over Europe. II: climate change scenarios (2071–2100). Climate Dynamics. 2004;23(7-8):839–58.
Lenoir J, Hattab T, Pierre G. Climatic microrefugia under anthropogenic climate change: implications for species redistribution. Ecography. 2017;40(2):253–66.
Benito GM, Robson TM, Hampe A. ΔTraitSDMs: species distribution models that account for local adaptation and phenotypic plasticity. New Phytologist. 2019;222:1757–65.
Pecchi M, Marchi M, Burton V, Giannetti F, Moriondo M, Bernetti I, et al. Species distribution modelling to support forest management. A literature review. Ecological Modelling. 2019;411:108817.
Oddou-Muratorio S, Davi H, Lefèvre F. Integrating evolutionary, demographic and ecophysiological processes to predict the adaptive dynamics of forest tree populations under global change. Tree Genetics & Genomes. 2020;16:67.
Sebald J, Thrippleton T, Rammer W, Bugmann H, Seidl R. Mixing tree species at different spatial scales: the effect of alpha, beta and gamma diversity on disturbance impacts under climate change. Journal of Applied Ecology. 2021;58:1749–63.
Alizoti PG, Kilimis K, Gallios P. Temporal and spatial variation of flowering among Pinus nigra Arn. clones under changing climatic conditions. Forest Ecology and Management. 2010;259(4):786–97.
Curtu AL, Alizoti P, Ballian D. Marginal/peripheral populations of forest tree species and their conservation status: report for southeastern Europe. Annals of Silvicultural Research. 2017;41(3):41–7.
Heer K, Behringer D, Piermattei A, Bässler C, Brandl R, Fady B, et al. Linking dendroecology and association genetics in natural populations: Stress responses archived in tree rings associate with SNP genotypes in silver fir (Abies alba Mill.). Molecular Ecology. 2018;27(6):1428–38.
••Aurelle D, Thomas S, Albert C, Bally M, Bondeau A, Boudouresque CF, et al. Biodiversity, climate change and adaptation in the Mediterranean. Ecosphere. 2021. https://doi.org/10.1002/ecs2.3915. A comprehensive review of adaptation risks and potentials in marine and terrestrial Mediterranean systems.
Matić S, Prpić B, Jakovac H, Vukelić J, Anić I, Delač D, editors. Forests of the Croatian Mediterranean: Croatian Academy of Forest Sciences; 2011. ISBN 978-953-985715-6 (https://www.sumari.hr/biblio/pdf/13223.pdf)
Pascual U, Muradian R, Brander L, Gómez-Maes JE, Paracchini MP, Zulian G, Alkemade R. Synergies and trade-offs between ecosystem service supply, biodiversity and habitat conservation status in Europe. Biological Conservation. 2012;1:155.
Morán-Ordóñez A, Ameztegui A, Cáceres D, de Miguel S, Lefèvre F, Brotons L, Coll L. Future trade-offs and synergies among ecosystem services in Mediterranean forests under global change scenarios. Ecosystem Services. 2020;45:101174.
Tufféry L, Davi H, López-García N, Rigolot E, Jean F, Stenger A, Lefèvre F. Adaptive measures for Mediterranean mountain forest ecosystem services under climate and land cover change in the Mont-Ventoux Natural Regional Park, France. Regional Environmental Change. 2021;21:12.
EUFORGEN. Forest genetic resources strategy for Europe: European Forest Institute; 2021.
Croitoru L. Valuing the non-timber forest products in the Mediterranean region. Ecological Economics. 2007;63(4):768–75.
Libiad M, Khabbach A, El Haissoufi M, Anestis I, Lamchouri F, Bourgou S, et al. Agro-alimentary potential of the neglected and underutilized local endemic plants of Crete (Greece), Rif-Mediterranean Coast of Morocco and Tunisia: perspectives and challenges. Plants. 2021;10:1770.
Sacchelli S, Borghi C, Fratini R, Bernetti I. Assessment and valorization of non-wood forest products in Europe: a quantitative literature review. Sustainability. 2021;13(6):3533.
Godet L, Devictor V. What conservation does. Trends in Ecology & Evolution. 2018;33(10):720–30.
Baciu GE, Dobrotă CE, Apostol EN. Valuing forest ecosystem services. Why Is an Integrative Approach Needed? Forests. 2021;12:677.
IPBES. The methodological assessment report on scenarios and models of biodiversity and ecosystem services. S. Ferrier, K. N. Ninan, P. Leadley, R. Alkemade, L. A. Acosta, H. R. Akcakaya, L. Brotons, W. W. L. Cheung, V. Christensen, K. A. Harhash, J. Kabubo-Mariara, C. Lundquist, M. Obersteiner, H. M. Pereira, G. Peterson, R. Pichs-Madruga, N. Ravindranath, C. Rondinini and B. A. Wintle (eds.). Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services, Bonn, Germany. 348 pages. 2016.
Raymond CM, Berry P, Breil M, Nita MR, Kabisch N, de Bel M, et al. An impact evaluation framework to support planning and evaluation of nature-based solutions projects. Report prepared by the EKLIPSE Expert Working Group on Nature-based Solutions to Promote Climate Resilience in Urban Areas. Wallingford, United Kingdom: Centre for Ecology & Hydrology; 2017.
Funding
We wish to acknowledge the following financial contributions:
- European Union’s Horizon 2020 research and innovation program under grant agreement No. 676876 “Optimizing the management and sustainable use of forest genetic resources in Europe” (GenTree);
- COST Action FP1202 “Strengthening conservation: a key issue for adaptation of marginal/peripheral populations of forest trees to climate change in Europe (MaP-FGR)”
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Disclaimer
The views expressed in this article are those of the author(s) and do not necessarily reflect the views or policies of the Food and Agriculture Organization of the United Nations.
Conflict of Interest
The authors declare no competing interests.
Human and Animal Rights and Informed Consent
This article does not contain any studies with human or animal subjects performed by any of the authors
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is part of the Topical Collection on Tree Genetics
The original online version of this article was revised due to a retrospective Open Access order and changes in authors’ affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fady, B., Esposito, E., Abulaila, K. et al. Forest Genetics Research in the Mediterranean Basin: Bibliometric Analysis, Knowledge Gaps, and Perspectives. Curr Forestry Rep 8, 277–298 (2022). https://doi.org/10.1007/s40725-022-00169-8
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40725-022-00169-8