

Japanese Knotweed Rhizome Bark Extract Inhibits Live SARS-CoV-2 In Vitro

1
Laboratory for Food Chemistry, Department of Analytical Chemistry, National Institute of Chemistry, Hajdrihova 19, 1000 Ljubljana, Slovenia
2
Veterinary Faculty, University of Ljubljana, Gerbičeva ulica 60, 1000 Ljubljana, Slovenia
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Bioengineering 2022, 9(9), 429; https://doi.org/10.3390/bioengineering9090429
Submission received: 20 July 2022
/
Revised: 12 August 2022
/
Accepted: 23 August 2022
/
Published: 1 September 2022
(This article belongs to the Special Issue Bioactive Formulations in Agri-Food-Pharma: Source and Applications, Volume II)
{kind=link}
{kind=link}
Abstract
:Coronavirus disease 2019 (COVID-19), a viral infectious respiratory disease, is caused by highly contagious severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and is responsible for the ongoing COVID-19 pandemic. Since very few drugs are known to be effective against SARS-CoV-2, there is a general need for new therapeutics, including plant-based drugs, for the prophylaxis and treatment of infections. In the current study, the activity of a 70% ethanolic(aq) extract of the rhizome bark of Japanese knotweed, an invasive alien plant species, was tested for the first time against the wild-type SARS-CoV-2 virus using a specific and robust virus neutralization test (VNT) on Vero-E6 cells, which best mimics the mechanism of real virus–host interaction. A statistically significant antiviral effect against SARS-CoV-2 (p-value < 0.05) was observed for the 50.8 µg mL−1 extract solution in cell medium. A suitable extract preparation was described to avoid loss of polyphenols throughout filtration of the extract, which was dissolved in cell medium containing fetal bovine serum (FBS). The significance of the differences between the sums of the test and control groups in the incidence of cytopathic effects (CPE) was determined using the one-way ANOVA test. A dose–response relationship was observed, with the cytotoxic effect occurring at higher concentrations of the extract (≥101.6 µg mL−1). The obtained results suggest possible use of this plant material for the production of various products (e.g., packaging, hygiene products, biodisinfectants, etc.) that would be useful against the spread of and for self-protection against COVID-19.
1. Introduction
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) [1] is a virus that is contagious to humans and some animal species and causes coronavirus disease 2019 (COVID-19), a viral respiratory illness responsible for the ongoing COVID-19 pandemic [2] declared by the World Health Organization on 11 March 2020 [3]. SARS-CoV-2 belongs to the Coronaviridae family, represented by enveloped viruses containing a positive-sense, single-stranded ribonucleic acid (+ssRNA) [4,5,6]. SARS-CoV-2 and other coronaviruses have four structural proteins known as spike, envelope, membrane, and nucleocapsid proteins. During viral infection, the spike protein promotes binding and fusion between the virus and the cell membrane [7]. The virus invades human cells by binding to the cell surface receptor angiotensin converting enzyme 2 (ACE2), a membrane glycoprotein [8,9]. The virus is transmitted from person to person mainly by close contact via aerosols and droplets from the respiratory air [10]. The nasal cavity respiratory epithelium is probably the predominant site of initial infection and virus replication [11]. Common symptoms and signs of a person infected with coronavirus include fever, cough, dyspnea, lymphopenia, abnormal results of chest computed tomography (CT), myalgia or fatigue, sputum production, headache, hemoptysis, diarrhea, etc. In more severe cases, the infection leads to acute respiratory distress syndrome, RNAemia, cardiac damage, secondary infections, and also death [12].
Depending on the target, there are two potential approaches to anti-coronavirus therapy, one targeting the coronavirus itself and the other targeting the support of the human immune system or human cells [7]. To date, few drugs are known to effectively inhibit SARS-CoV-2. Among specific drugs against SARS-CoV-2, Veklury (remdesivir) and Olumiant (baricitinib) have been approved by the FDA [13]. The EMA has approved several treatments for COVID-19: Evusheld (tixagevimab/cilgavimab), Kineret (anakinra), Paxlovid (PF-07321332/ritonavir), Regkirona (regdanvimab), RoActemra (tocilizumab), Ronapreve (casirivimab/imdevimab), Veklury (remdesivir), and Xevudy (sotrovimab) [14].
Plant-based drugs, including Japanese knotweed, have been used to treat various viral infections. The identification of phytochemicals with health-beneficial effects would be crucial for the development of new drugs against SARS-CoV-2 infection.
Japanese knotweed (Fallopia japonica Houtt., Reynoutria japonica Houtt., Polygonum cuspidatum Siebold & Zucc.) is an alien plant species native to East Asia that has become invasive in Europe and North America [15]. The rhizome extracts of Japanese knotweed have been tested in various biological studies [16], and the extracts themselves or their compounds showed antioxidant [17,18,19,20,21,22,23,24], antiproliferative [17], estrogenic [25], antiatherosclerotic [26], anti-inflammatory [27], antibacterial [28], and antiviral activities. Japanese knotweed ethanolic extract inhibits the lytic cycle and reduces the production of Epstein –Barr (EBV) viral particles [29]. Ethanolic and water extracts (the latter at a higher dose) inhibit the production of hepatitis B virus (HBV) [30]. Methanolic extract inhibits infection with the mosquito-borne pathogen dengue virus (DENV) in the early viral entry phases and also reduces the infectivity of hepatitis C virus (HCV) and Zika virus (ZIKV) [31]. The antiviral effect of Japanese knotweed water extract was investigated in a model of acquired immunodeficiency syndrome in mice, and immunodeficiency was partially inhibited [32]. In addition, a 70% ethanolic(aq) extract was tested against human immunodeficiency virus type 1 (HIV-1) and inhibited HIV-1-induced syncytium formation [33]. Bioactivity-guided fractionation enabled the isolation of 20 phenolic compounds with anti-HIV potential, and potent antiviral activity against HIV-1 was demonstrated for resveratrol, (+)-catechin, emodin-8-O-glucoside, and 5,7-dimethoxyphthalide [33]. Japanese knotweed water extract and its bioactive components resveratrol and emodin inhibit the replication of influenza A (H1N1) virus via interference with the mechanisms of the Toll-like receptor-9 pathway [34]. Furthermore, bioactivity-guided fractionation of the ethyl acetate extract was performed, and of the seven compounds isolated, resveratrol, (E)-3,5,12-trihydroxystilbene-3-O-beta-D-glucopyranoside-2′-(3″,4″,5″-trihydroxybenzoate) and catechin-3-O-gallate showed an inhibitory effect on neuraminidase activity, while the last two compounds also showed inhibitory activity against H1N1 [35].
Two important substances of Japanese knotweed rhizomes with well-studied and confirmed antiviral activity are emodin and resveratrol [36,37,38,39,40,41,42,43,44,45], phenolic compounds belonging to the anthraquinone and stilbene groups, respectively.
For emodin extracted from the rhizome of Japanese knotweed, antiviral activity against human simplex virus type 1 (HSV-1) was observed in guinea pigs [36]. Emodin inhibits the DNA replication of HBV [44]. Emodin and an ethyl acetate subfraction of Japanese knotweed rhizome, which contained 68.2% emodin, inhibited the expression of EBV immediate-early proteins and DNA replication [43]. Emodin isolated from Japanese knotweed inhibited the entry and replication of Coxsackie B4 virus (CVB4) and improved the survival of infected mice when administered orally [45].
Resveratrol inhibited the induced expression of early EBV antigen in Raji cells [41] and was proven useful in preventing the proliferation of EBV [37]. In addition, resveratrol inhibited the replication of human cytomegalovirus (HCMV) [38], varicella-zoster virus (VZV) [40], HSV-1 [39], and even HIV-1 [42].
Due to the beneficial effects of some traditional Chinese medicine (TCM) plants on SARS-CoV virus infection and other coronaviruses, these plants and their secondary metabolites have attracted attention and have been studied also within the current pandemic [46].
Emodin inhibited the interaction of SARS-CoV spike protein and ACE2 [47] and blocked coronavirus SARS-CoV and HCoV-OC43 ion channel 3a, which impaired virus release [48].
Resveratrol inhibited Middle East respiratory syndrome coronavirus (MERS-CoV) infection and prolonged cellular survival after infection. The expression of nucleocapsid protein essential for MERS-CoV replication decreased, and apoptosis induced by MERS-CoV in vitro was downregulated, after resveratrol treatment [49].
The aim of the current study was to test the antiviral activity of the 70% ethanol(aq) extract of Japanese knotweed rhizome bark extract against live SARS-CoV-2 in vitro for the first time. A search in the literature shows a lack of in vitro studies as opposed to in silico studies to test the activity of this plant extract; the exceptions are Nawrot-Hadzik et al., 2021 [50], reporting the in vitro SARS-CoV-2 Mpro enzyme inhibitory activity of 70% acetone extract of the plant whole rhizome and Lin et al., 2022 [51], reporting the blocked entry of the SARS-CoV-2 pseudotyped virus into fibroblasts by water and ethanol extracts of the rhizome and root. The studies discussing Japanese knotweed or its compounds in connection with SARS-CoV are elaborated in detail in Results and Discussion.
2. Materials and Methods
2.1. Preparation of the Japanese Knotweed Rhizome Bark Extract
Rhizomes of Japanese knotweed were harvested in Ljubljana, Slovenia (Vrhovci, Mali Graben river bank; N 46°02′33.9″, E 14°27′00.9″). A voucher specimen was deposited in the Herbarium LJU (LJU10143477). The rhizomes were cleaned with tap water, the bark was peeled and lyophilized at −50 °C for 24 h (Micro Modulyo, IMAEdwards, Bologna, Italy), and the obtained dry material was frozen with liquid N2 and pulverized by a Mikro-Dismembrator S (Sartorius, Goettingen, Germany; 1 min, 1700 min−1). The rhizome bark powder (1 g) was extracted with 20 mL of 70% ethanol(aq) (ethanol absolute anhydrous was purchased from Carlo Erba Reagents (Val de Reuil, France), and a Milli-Q water purification system (18 MΩ cm−1; Millipore, Bedford, MA, USA) was used to obtain ultrapure water). Vortexing (5 min), sonication (15 min) and centrifugation (5 min, 6700× g) were executed and the supernatant was transferred to a pre-weighted vial, while the extraction of the solid residue was repeated with 10 mL 70% ethanol(aq). The solvent of the pooled supernatants was evaporated under N2 flow. The dry extract (414.17 mg) was dissolved in 127.37 mL of cell medium (extract stock solution) in a sterile hood and was used for antiviral activity assays without filtration.
2.2. Native SARS-CoV-2 Virus Neutralization Test (VNT)
Different concentrations of the extract from the stock solution (concentration 3.25 mg mL−1) in the culture media (ATCC; E-MEM with addition of 10% of FBS and 1% of standard antibiotic and antimycotic (Gibco, Grand Island, NY, USA; Anti-Anti 100x) were prepared in volume ratios of 1:64, 1:128, and 1:256. The number of infectious viral particles was quantified using the Median Tissue Culture Infectious Dose (TCID50) test. The assay is based on adding a serial dilution of the virus sample to the susceptible cells in a 96-well plate. The dilution at which 50% of the wells show CPE is used to mathematically calculate the TCID50 of the virus sample as generally described.
Extract dilutions (25 μL) and medium (25 μL) containing 6.2 TCID50 SARS-CoV-2 as virus working concentrations (SARS-CoV-2, 4265/20; EVAg) were mixed and incubated at 37 °C and 5% CO2 for 7 h. Each dilution of the extract and virus was incubated in 48 replicates in 96-well tissue culture plates (tissue culture test-plate 96F, TPP Techno Plastic Products AG, Trasadingen, Swiss). After the incubation period, 3.5 × 105 Vero-E6 (African green monkey kidney cell line; ATCC CRL-1587) cell suspension was added in a volume of 100 μL to reach a final volume of 150 μL/well. To standardize the test procedures, a positive control with a working dilution of virus that was not incubated with the extract under the same conditions and volumes was included. After 110 h, the microplates were observed by inverted light microscope (ECLIPSE Ts2R, NIKON Instruments, Melville, NY, USA) at 400-fold magnification under a microscope equipped with an LED display. Characteristic morphological changes in the cells (rounding of adherent infected cells) in the culture that were the target of the SARS-CoV-2 replication were labelled as CPE. The other cellular transformation (cell dissolution and loss of cellular ultrastructure) was declared as a cytotoxic, necrotic effect. Each well in which the CPE was detected was labelled as positive for virus replication.
Before the main experiment, combinations of screening tests with different virus concentrations (virus working solution 100 TCID50 and subsequent dilutions in volume ratios of 1:2, 1:4, 1:8, 1:16, 1:32, 1:64, and 1:128) were performed in combination with different concentrations of extract (dilutions of stock solution in volume ratios of 1:2, 1:4, 1:8, 1:16, 1:32, 1:64, 1:128, 1:256, 1:512, 1:1024, and 1:2048) to determine the optimal concentration range for determining the cytotoxic properties of the extract and its potential antiviral activity under the same conditions as those under which the main experiment was later performed.
2.3. Statistical Analysis
The significance of the differences between the sums of the test and control groups in CPE incidence was determined using the one-way ANOVA test. A p-value of <0.05 was considered statistically significant. All the data were analyzed using Microsoft Excel 2016 (v16.0, Microsoft, Redmond, WA, USA).
3. Results and Discussion
A green solvent, 70% ethanol(aq), suitable for tincture preparation was selected as an extraction medium for Japanese knotweed rhizome bark due to the obtained higher extraction yield (44.3%) in comparison to other solvents [52]. It is also considered less harmful when present as a residual solvent in pharmaceutical formulations compared to other organic solvents [53]. An additional advantage of ethanol is its commercial availability as a food-grade solvent. Japanese knotweed rhizome bark 70% ethanolic(aq) extract was tested for its antioxidant activity in our previous study and was found to possess potent and stable time-dependent antioxidant activity [52].
In the search for an herbal antiviral drug and supported by already proven antiviral properties against other viruses, Japanese knotweed is one of several candidates to be tested against SARS-CoV-2. Within the current study we investigated the effect of the 70% ethanol(aq) Japanese knotweed rhizome bark extract on the antiviral properties of a wild-type SARS-CoV-2 using the virus neutralization test (VNT), which is routinely used in laboratory diagnostics of viral diseases or in the detection of specific antibodies [54]. The test is also a suitable research tool for studying all potential antiviral substances, such as antiviral peptides, drugs, or disinfectants. The test detects prevention or inhibition of cytopathic effect (CPE) and viral replication if neutralizing antibodies or substances with antiviral properties are detected [54].
In the current study, the extract was incubated together with SARS-CoV-2 for 7 h before cell exposure.
It is illusory to expect a neutralizing effect in the range of the specific neutralizing antibodies (NAbs), making the partial reduction in the cytopathic effect in such an experimental model a realistic expectation.
One of the biological effects of NAbs is to reduce the infectivity of the virus by their specific binding to viral ligands, which block the attachment of the virus to the cell receptor, which is the principle of virus neutralization [54]. Other antiviral mechanisms include a wide spectrum of possibly undefined interactions at the molecular level between the virus and the host cell, leading to impaired virus infectivity or even to the destruction of the virus. The gold standard for evaluating the in vitro neutralization ability or antiviral properties of substances is the virus neutralization test with native, infectious virus, which most closely mimics the mechanism of virus–host interaction in natural infection. Accuracy is its key feature, requiring manipulation of wild-type live virus, and is therefore performed in level III biosafety standards laboratories. The closest alternative to this type of testing is the pseudovirus neutralization test, which uses a noninfectious pseudovirus and is therefore a safer test performed in level II laboratories, but the results must be interpreted with a certain degree of caution [54].
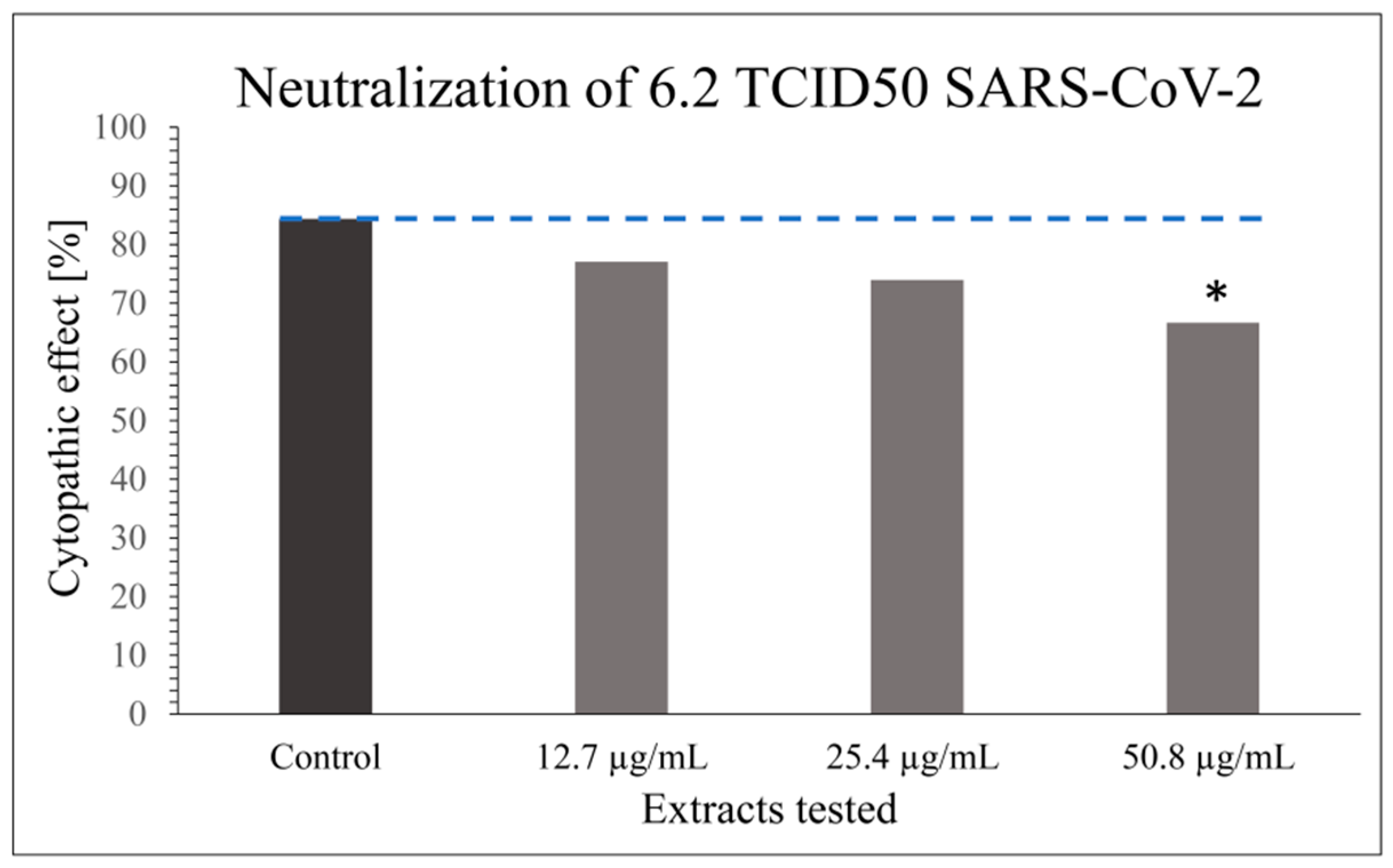
Modifications of VNT have been described in the scientific literature to investigate the antiviral properties of such herbal extracts [51]. However, only laboratory testing models with a wild-type virus can most suitably mimic the natural viral infection of the host by allowing long-term virus–cell contact with prolonged antiviral contact of the virus extract in the experiment. The obtained results indicate that the tested extract has a certain level of inhibitory properties on the viral infectivity cycle; however, these are not as potent as the specific neutralizing antibody. CPE or antiviral effect of SARS-CoV-2 was studied in 48 parallels at each virus’ and extract’s concentration, taking into consideration only dilutions of the extract stock solution that did not cause cytotoxicity (1:64; 1:128; 1:256; v/v) in combination with the optimal working concentration of the virus (6.2 TCID50 SARS-CoV-2) (Figure 1 and Figure 2) as previously determined by screening tests.
A statistically significant antiviral effect (reduction in CPE) against SARS-CoV-2 was observed for the stock solution diluted 1:64 (v/v) (i.e., 50.8 µg mL−1) compared to the incidence of CPE in the positive control, where the virus-working concentration was inoculated on Vero-E6 cells only; p < 0.05 (Figure 1). Besides the statistically significant result for the CPE reduction effect of the extract at 50.8 µg mL−1 compared to the positive control, a dose–response relationship was generally noticed for all the extract concentrations (Figure 1).
The interactions between Japanese knotweed flavan-3-ols and proanthocyanidins [52,55] and FBS albumin in the cell medium should be considered when evaluating Japanese knotweed extract using in vitro cell culture systems. Namely, among various biological activities, Japanese knotweed polyphenols possess astringent activity [56]. The VNT executed with the filtered extract dissolved in cell medium did not show promising results. Therefore, final experiments were performed with the non-filtered extract to avoid possible removal of these complexes. According to the literature, interactions between flavan-3-ols and FBS albumin may induce cytotoxicity at high concentrations of flavan-3-ols [57]. In the current study, the cytotoxic effect was observed at extract concentrations above 50.8 µg mL−1. Although SARS-CoV-2 inhibitory activity was expected to be even higher at higher concentrations of the extract, this could not be evaluated at cytotoxic concentrations.
The mechanisms of antiviral activity on SARS-CoV-2 inhibitory activity and the main antiviral compounds of the examined extract as well as possible cell protection effect should be investigated on a molecular level in a further study. To support the obtained result, a few studies elaborating the effect of Japanese knotweed extract and its compounds on SARS-CoV virus are described below; however, none of these studies involve live, wild-type SARS-CoV-2 virus.
An herbal drug called Shufeng Jiedu, consisting of eight medicinal plants including Japanese knotweed, is known for its antiviral, anti-inflammatory, and immunomodulatory effects in acute lung diseases and was discussed as a promising candidate for the treatment of COVID-19 [58]. However, the antiviral and anti-inflammatory effects of Shufeng Jiedu were tested in a HCoV-229E (group 1 coronavirus) mouse model [58]. The formulation reduced viral load and decreased inflammatory factors in the lungs while increasing the concentration of CD4+ and CD8+ cells in the blood [58]. Direct binding of polydatin (an anthraquinone from the Japanese knotweed rhizome [52,55]), quercetin, and wogonin to the major protease of SARS-CoV-2 was demonstrated in silico [58]. Clinical data for Shufeng Jiedu added to standard antiviral therapy showed significant reduction in the COVID-19 clinical recovery time. Still, largescale, randomized, placebo-controlled, double-blinded clinical trials are lacking.
In addition, Baidu Jieduan granules containing Japanese knotweed and eleven other herbs have been clinically evaluated for efficacy and safety in the treatment of moderate COVID-19 [59]. Nonetheless, placebo-controlled and double-blinded clinical trial design is lacking.
The results of network pharmacology and bioinformatics analysis suggest that Japanese knotweed material is a promising therapeutic agent against COVID-19 [60].
The antiviral activity of purchased standards of polydatin and resveratrol, typical compounds of Japanese knotweed, has been demonstrated in vitro against HCoV-OC43 strain, an alternative model for SARS-CoV-2 [61]. Furthermore, computer-aided virtual screening was used to predict the binding site, and surface plasmon resonance (SPR) analysis was employed to confirm the interaction. SPR results showed a specific affinity of polydatin and resveratrol toward SARS-CoV 3CLpro and PLpro proteins as well as toward SARS-CoV-2. Moreover, Japanese knotweed was suggested as a potential therapeutic agent for pulmonary fibrosis caused by COVID-19 based on network pharmacology and data mining [62].
In other studies, a decreased infectivity of the SARS-CoV-2 spike pseudovirus in HEK293T-ACE2 cells was observed by polydatin [63], while resveratrol-inhibited SARS-CoV-2 replication (0–99.3%) in a dose-dependent manner (0–25 µM) with an EC90 and EC50 (50% and 90% maximal effective concentration, respectively) 11.42 and 10.66 µM, respectively. Higher concentrations of resveratrol (>50 µM) posed cytotoxicity on Vero-E6 cells [64].
In the study by Nawrot-Hadzik et al., 2021 [50], 25 compounds known to be present in the rhizomes of Reynoutria japonica (Japanese knotweed) and Reynoutria sachalinensis were docked into the main protease binding site of SARS-CoV-2. Further, 11 of them together with the extracts of both plants were tested in vitro for inhibition of SARS-CoV-2 Mpro enzyme [50]. Vanicosides A and B, isolated from the rhizomes of Reynoutria sachalinensis, showed moderate inhibitory activity against SARS-CoV-2 Mpro, while acetone extract and especially the butanol fractions of plants (of tested: dichloromethane, diethyl ether, ethyl acetate, n-butanol, and water fractions) containing vanicosides and polymerized procyanidins showed strong inhibition against SARS-CoV-2 Mpro [50]. Water and 90% ethanol(aq) extracts of Japanese knotweed rhizome and root blocked the entry of SARS-CoV-2 pseudotyped virus into HEK293T-ACE2 cells and zebrafish larvae and were shown to inhibit the spike protein–ACE2 receptor interaction and 3CL protease activity. Anti-SARS-CoV-2 activity was confirmed for the extract component gallic acid [51].
Some doubts and counter-opinions about the antiviral activity of Japanese knotweed against SARS-CoV-2 are also present in the literature. For instance, although the frequency of COVID-19 infection cases should be lower in Asian countries where Japanese knotweed occurs as a component of local foods, this has not been the case [65].
The results of the current study could be prospectively used to formulate or at least promote food supplements containing a Japanese knotweed rhizome bark extract for the prevention or complementary treatment of COVID-19 patients. A biofoil enriched with this extract which could be used as active packaging for food, drugs, and cosmetics to protect food or other packed contents from bacteria and oxidation has already been formulated by our group [66]. These foils may potentially prevent the spread of coronavirus (or other virus species), since another possible way for coronavirus to spread, besides through respiratory air (as mentioned in the introduction) is via indirect contact with contaminated surfaces. The virus remains viable and infectious in aerosols for hours and on surfaces for up to a day [67]. It is particularly stable on conventional plastic and stainless steel [67]. Other hygiene products such as soaps (soap destabilizes the lipid bilayer of viruses [68]) and disinfectant solutions (ethanol or isopropanol inactivate human coronavirus [69]) could also be enriched with Japanese knotweed rhizome bark extract to enhance their effect (act as bio-disinfectants). In this way, we would fight two battles at the same time: potential new COVID-19 waves and the invasiveness of Japanese knotweed, as the best way to eradicate or at least restrict this plant from Europe and North America (where it represents a major economic and environmental problem) is via mechanical excavation, which, on the other hand, is a necessary step to obtain extracts from its rhizome bark.
Author Contributions
Conceptualization, U.J., K.N. and T.M.; methodology, K.N., U.J. and T.M.; formal analysis, T.M.; investigation, U.J., K.N. and T.M.; resources, K.N. and T.M.; data curation, K.N. and U.J.; writing—original draft preparation, U.J., K.N. and T.M.; writing—review and editing, K.N. and U.J.; visualization, T.M. and U.J.; funding acquisition, K.N. and T.M. All authors have read and agreed to the published version of the manuscript.
Funding
The study was carried out with financial support from the Slovenian Research Agency (P1-0005 and P4-0092).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data generated for this study are included in the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gorbalenya, A.E.; Baker, S.C.; Baric, R.S.; De Groot, R.J.; Drosten, C.; Gulyaeva, A.A.; Haagmans, B.L.; Lauber, C.; Leontovich, A.M.; Neuman, B.W.; et al. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef]
- COVID-19: Epidemiology, Virology, and Prevention. Available online: https://www.uptodate.com/contents/covid-19-epidemiology-virology-and-prevention (accessed on 14 November 2021).
- WHO Director-General’s opening remarks at the media briefing on COVID-19—11 March 2020. World Health Organization (WHO) (Press Release). Available online: https://www.who.int/director-general/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19---11-march-2020 (accessed on 14 November 2021).
- Cui, J.; Li, F.; Shi, Z.L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Machhi, J.; Herskovitz, J.; Senan, A.M.; Dutta, D.; Nath, B.; Oleynikov, M.D.; Blomberg, W.R.; Meigs, D.D.; Hasan, M.; Patel, M.; et al. The natural history, pathobiology, and clinical manifestations of SARS-CoV-2 infections. J. Neuroimmune Pharmacol. 2020, 15, 359–386. [Google Scholar] [CrossRef]
- Pal, M.; Berhanu, G.; Desalegn, C.; Kandi, V. Severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2): An update. Cureus 2020, 12, e7423. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Liu, Y.; Yang, Y.; Zhang, P.; Zhong, W.; Wang, Y.; Wang, Q.; Xu, Y.; Li, M.; Li, X.; et al. Analysis of therapeutic targets for SARS-CoV-2 and discovery of potential drugs by computational methods. Acta Pharm. Sin. B 2020, 10, 766–788. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef]
- Coronavirus Disease (COVID-19): How Is It Transmitted? Available online: https://web.archive.org/web/20201015230546/https://www.who.int/news-room/q-a-detail/coronavirus-disease-covid-19-how-is-it-transmitted (accessed on 14 November 2021).
- Hou, Y.J.; Okuda, K.; Edwards, C.E.; Martinez, D.R.; Asakura, T.; Dinnon, K.H., 3rd; Kato, T.; Lee, R.E.; Yount, B.L.; Mascenik, T.M.; et al. SARS-CoV-2 Reverse genetics reveals a variable infection gradient in the respiratory tract. Cell 2020, 182, 429–446. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Coronavirus (COVID-19), Drugs. Available online: https://www.fda.gov/drugs/emergency-preparedness-drugs/coronavirus-covid-19-drugs (accessed on 12 August 2022).
- COVID-19 Treatments, European Medicines Agency. Available online: https://www.ema.europa.eu/en/human-regulatory/overview/public-health-threats/coronavirus-disease-covid-19/treatments-vaccines/covid-19-treatments (accessed on 12 August 2022).
- Balogh, L. Japanese, Giant and Bohemian knotweed. In The Most Important Invasive Plants in Hungary; Botta-Dukat, Z., Balogh, L., Eds.; HAS Institute of Ecology and Botany: Budapest, Hungary, 2008; pp. 13–33. [Google Scholar]
- Zhang, H.; Li, C.; Kwok, S.-T.; Zhang, Q.-W.; Chan, S.-W. A review of the pharmacological effects of the dried root of Polygonum cuspidatum (Hu Zhang) and its constituents. Evid.-Based Complementary Altern. Med. 2013, 2013, 208349. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-W.; Yang, F.-J.; Chen, C.-L.; Lee, W.-T.; Chen, R.-S. Free radical scavenging activity and antiproliferative potential of Polygonum cuspidatum root extracts. J. Nat. Med. 2010, 64, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-Y.; Chan, Y.-P.; Chang, J. Antioxidant activity of extract from Polygonum cuspidatum. Biol. Res. 2007, 40, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Pogačnik, L.; Rogelj, A.; Ulrih, N.P. Chemiluminescence method for evaluation of antioxidant capacities of different invasive knotweed species. Anal. Lett. 2015, 49, 350–363. [Google Scholar] [CrossRef]
- Lachowicz, S.; Oszmianski, J. Profile of bioactive compounds in the morphological parts of wild Fallopia japonica (Houtt) and Fallopia sachalinensis (F. Schmidt) and their antioxidative activity. Molecules 2019, 24, 1436. [Google Scholar] [CrossRef]
- Ardelean, F.; Moacă, E.A.; Păcurariu, C.; Antal, D.S.; Dehelean, C.; Toma, C.-C.; Drăgan, S. Invasive Polygonum cuspidatum: Physico-chemical analysis of a plant extract with pharmaceutical potential. Studia Univ. Vasile Goldis Arad Ser. Stiintele Vietii 2016, 26, 415–421. [Google Scholar]
- Kurita, S.; Kashiwagi, T.; Ebisu, T.; Shimamura, T.; Ukeda, H. Content of resveratrol and glycoside and its contribution to the antioxidative capacity of Polygonum cuspidatum (Itadori) harvested in Kochi. Biosci. Biotechnol. Biochem. 2014, 78, 499–502. [Google Scholar] [CrossRef]
- Chan, C.-L.; Gan, R.-Y.; Corke, H. The phenolic composition and antioxidant capacity of soluble and bound extracts in selected dietary spices and medicinal herbs. Int. J. Food Sci. Technol. 2016, 51, 565–573. [Google Scholar] [CrossRef]
- Nawrot-Hadzik, I.; Ślusarczyk, S.; Granica, S.; Hadzik, J.; Matkowski, A. Phytochemical diversity in rhizomes of three Reynoutria species and their antioxidant activity correlations elucidated by LC-ESI-MS/MS analysis. Molecules 2019, 24, 1136. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, X.; Zhang, Y.; Xu, Q.; Xiao, H.; Liang, X. Analysis of estrogenic compounds in Polygonum cuspidatum by bioassay and high performance liquid chromatography. J. Ethnopharmacol. 2006, 105, 223–228. [Google Scholar] [CrossRef]
- Xue, Y.; Liang, J. Screening of bioactive compounds in rhizoma Polygoni cuspidati with hepatocyte membranes by HPLC and LC-MS. J. Sep. Sci. 2014, 37, 250–256. [Google Scholar] [CrossRef]
- Fan, P.; Zhang, T.; Hostettmann, K. Anti-inflammatory activity of the invasive neophyte Polygonum cuspidatum Sieb. and Zucc. (Polygonaceae) and the chemical comparison of the invasive and native varieties with regard to resveratrol. J. Tradit. Complementary Med. 2013, 3, 182–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, B.; Cai, Y.-Z.; Brooks, J.D.; Corke, H. Antibacterial properties of Polygonum cuspidatum roots and their major bioactive constituents. Food Chem. 2008, 109, 530–537. [Google Scholar] [CrossRef]
- Yiu, C.-Y.; Chen, S.-Y.; Huang, C.-W.; Yeh, D.-B.; Lin, T.-P. Inhibitory effects of Polygonum cuspidatum on the Epstein-Barr virus lytic cycle. J. Food Drug Anal. 2011, 19, 107–113. [Google Scholar] [CrossRef]
- Chang, J.-S.; Liu, H.-W.; Wang, K.-C.; Chen, M.-C.; Chiang, L.-C.; Hua, Y.-C.; Lin, C.-C. Ethanol extract of Polygonum cuspidatum inhibits hepatitis B virus in a stable HBV-producing cell line. Antivir. Res. 2005, 66, 29–34. [Google Scholar] [CrossRef]
- Kuo, Y.-T.; Liu, C.-H.; Li, J.-W.; Lin, C.-J.; Jassey, A.; Wu, H.-N.; Perng, G.C.; Yen, M.-H.; Lin, L.T. Identification of the phytobioactive Polygonum cuspidatum as an antiviral source for restricting dengue virus entry. Sci. Rep. 2020, 10, 16378. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.I.; Hong-Xia, W.A.; Zuo-Yi, B.A.; Guan-Fu, Z.H. Evaluation of antiviral effect of Polygonum cuspidatum water extract with a model of murine acquired immunodeficiency syndrome. Virol. Sin. 1998, 13, 311. [Google Scholar]
- Lin, H.-W.; Sun, M.-X.; Wang, Y.-H.; Yang, L.-M.; Yang, Y.-R.; Huang, N.; Xuan, L.-J.; Xu, Y.-M.; Bai, D.-L.; Zheng, Y.-T.; et al. Anti-HIV activities of the compounds isolated from Polygonum cuspidatum and Polygonum multiflorum. Planta Med. 2010, 76, 889–892. [Google Scholar] [CrossRef]
- Lin, C.J.; Lin, H.J.; Chen, T.H.; Hsu, Y.A.; Liu, C.S.; Hwang, G.Y.; Wan, L. Polygonum cuspidatum and its active components inhibit replication of the influenza virus through toll-like receptor 9-induced interferon beta expression. PLoS ONE 2015, 10, e0117602. [Google Scholar] [CrossRef]
- Chen, K.-T.; Zhou, W.-L.; Liu, J.-W.; Zu, M.; He, Z.N.; Du, G.H.; Chen, W.W.; Liu, A.L. Active neuraminidase constituents of Polygonum cuspidatum against influenza A(H1N1) influenza virus. Zhongguo Zhong Yao Za Zhi 2012, 37, 3068–3073. [Google Scholar] [CrossRef]
- Wang, Z.; Huang, T.; Guo, S.; Wang, R. Effects of emodin extracted from Rhizoma Polygoni Cuspidati in treating HSV-1 cutaneous infection in guinea pigs. J. Anhui Tradit. Chin. Med. Coll. 2003, 22, 36–38. [Google Scholar]
- Yiu, C.-Y.; Chen, S.-Y.; Chang, L.-K.; Chiu, Y.-F.; Lin, T.-P. Inhibitory effects of resveratrol on the Epstein-Barr virus lytic cycle. Molecules 2010, 15, 7115–7124. [Google Scholar] [CrossRef] [PubMed]
- Evers, D.L.; Wang, X.; Huong, S.-M.; Huang, D.Y.; Huang, E.-S. 3,4′,5-Trihydroxy-trans-stilbene (resveratrol) inhibits human cytomegalovirus replication and virus-induced cellular signaling. Antivir. Res. 2004, 63, 85–95. [Google Scholar] [CrossRef]
- Docherty, J.J.; Fu, M.M.; Stiffler, B.S.; Limperos, R.J.; Pokabla, C.M.; DeLucia, A.L. Resveratrol inhibition of herpes simplex virus replication. Antivir. Res. 1999, 43, 145–155. [Google Scholar] [CrossRef]
- Docherty, J.J.; Sweet, T.J.; Bailey, E.; Faith, S.A.; Booth, T. Resveratrol inhibition of varicella-zoster virus replication in vitro. Antivir. Res. 2006, 72, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Kapadia, G.J.; Azuine, M.A.; Tokuda, H.; Takasaki, M.; Mukainaka, T.; Konoshima, T.; Nishino, H. Chemopreventive effect of resveratrol, sesamol, sesame oil and sunflower oil in the Epstein-Barr virus early antigen activation assay and the mouse skin two-stage carcinogenesis. Pharmacol. Res. 2002, 45, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-S.; Zhou, Y.; Wu, M.-R.; Zhou, H.-S.; Xu, F. Resveratrol inhibited Tat-induced HIV-1 LTR transactivation via NAD+-dependent SIRT1 activity. Life Sci. 2009, 85, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Yiu, C.-Y.; Chen, S.-Y.; Yang, T.-H.; Chang, C.-J.; Yeh, D.-B.; Chen, Y.-J.; Lin, T.-P. Inhibition of Epstein-Barr virus lytic cycle by an ethyl acetate subfraction separated from Polygonum cuspidatum root and its major component, emodin. Molecules 2014, 19, 1258–1272. [Google Scholar] [CrossRef]
- Shuang-Suo, D.; Zhengguo, Z.; Yunru, C.; Xin, Z.; Baofeng, W.; Lichao, Y.; Yan’an, C. Inhibition of the replication of hepatitis B virus in vitro by emodin. Med. Sci. Monit. 2006, 12, 302–306. [Google Scholar]
- Liu, Z.; Wei, F.; Chen, L.-J.; Xiong, H.-R.; Liu, Y.-Y.; Luo, F.; Hou, W.; Xiao, H.; Yang, Z.-Q. In vitro and in vivo studies of the inhibitory effects of emodin isolated from Polygonum cuspidatum on Coxsakievirus B4. Molecules 2013, 18, 11842–11858. [Google Scholar] [CrossRef]
- Yang, Y.; Islam, M.S.; Wang, J.; Li, Y.; Chen, X. Traditional Chinese medicine in the treatment of patients infected with 2019-new coronavirus (SARS-CoV-2): A review and perspective. Int. J. Biol. Sci. 2020, 16, 1708–1717. [Google Scholar] [CrossRef]
- Ho, T.-Y.; Wu, S.-L.; Chen, J.-C.; Li, C.-C.; Hsiang, C.-Y. Emodin blocks the SARS coronavirus spike protein and angiotensin-converting enzyme 2 interaction. Antivir. Res. 2007, 74, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Wang, K.; Yu, W.J.; Sun, B.; Schwarz, W. Emodin inhibits current through SARS-associated coronavirus 3a protein. Antivir. Res. 2011, 90, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-C.; Ho, C.-T.; Chuo, W.-H.; Li, S.; Wang, T.T.; Lin, C.-C. Effective inhibition of MERS-CoV infection by resveratrol. BMC Infect. Dis. 2017, 17, 144. [Google Scholar] [CrossRef] [PubMed]
- Nawrot-Hadzik, I.; Zmudzinski, M.; Matkowski, A.; Preissner, R.; Kęsik-Brodacka, M.; Hadzik, J.; Drag, M.; Abel, R. Reynoutria rhizomes as a natural source of SARS-CoV-2 Mpro inhibitors-molecular docking and in vitro study. Pharmaceuticals 2021, 14, 742. [Google Scholar] [CrossRef]
- Lin, S.; Wang, X.; Tang, R.W.; Lee, H.C.; Chan, H.H.; Choi, S.S.A.; Dong, T.T.; Leung, K.W.; Webb, S.E.; Miller, A.L.; et al. The extracts of Polygonum cuspidatum root and rhizome block the entry of SARS-CoV-2 wild-type and omicron pseudotyped viruses via inhibition of the S-protein and 3CL protease. Molecules 2022, 27, 3806. [Google Scholar] [CrossRef]
- Jug, U.; Naumoska, K.; Vovk, I. (−)-Epicatechin—An important contributor to the antioxidant activity of Japanese knotweed rhizome bark extract as determined by antioxidant activity-guided fractionation. Antioxidants 2021, 10, 133. [Google Scholar] [CrossRef]
- Grodowska, K.; Parczewski, A. Organic solvents in the pharmaceutical industry. Acta Pol. Pharm. 2010, 67, 3–12. Available online: https://www.ptfarm.pl/wydawnictwa/czasopisma/acta-poloniae-pharmaceutica/110/-/12992 (accessed on 10 September 2020).
- Lu, Y.; Wang, J.; Li, Q.; Hu, H.; Lu, J.; Chen, Z. Advances in neutralization assays for SARS-CoV-2. Scand. J. Immunol. 2021, 94, e13088. [Google Scholar] [CrossRef]
- Jug, U.; Glavnik, V.; Vovk, I.; Makuc, D.; Naumoska, K. Off-line multidimensional high performance thin-layer chromatography for fractionation of Japanese knotweed rhizome bark extract and isolation of flavan-3-ols, proanthocyanidins and anthraquinones. J. Chromatogr. A 2021, 1637, 461802. [Google Scholar] [CrossRef]
- Cos, P.; De Bruyne, T.; Hermans, N.; Apers, S.; Vanden Berge, D.; Vlietinck, A.J. Proanthocyanidins in health care: Current and new trends. Curr. Med. Chem. 2004, 11, 1345–1359. [Google Scholar] [CrossRef]
- Fujii, Y.; Suhara, Y.; Sukikara, Y.; Teshima, T.; Hirota, Y.; Yoshimura, K.; Osakabe, N. Elucidation of the interaction between flavan-3-ols and bovine serum albumin and its effect on their in-vitro cytotoxicity. Molecules 2019, 24, 3667. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Shi, Y.; Su, J.; Friedemann, T.; Tao, Z.; Lu, Y.; Ling, Y.; Lv, Y.; Zhao, R.; Geng, Z.; et al. Shufeng Jiedu, a promising herbal therapy for moderate COVID-19: Antiviral and anti-inflammatory properties, pathways of bioactive compounds, and a clinical real-world pragmatic study. Phytomedicine 2021, 85, 153390. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xie, Q.; Xu, X.; Sun, S.; Fan, T.; Wu, X.; Qu, Y.; Che, J.; Huang, T.; Li, H.; et al. Baidu Jieduan granules, traditional Chinese medicine, in the treatment of moderate coronavirus disease-2019 (COVID-19): Study protocol for an open-label, randomized controlled clinical trial. Trials 2021, 22, 476. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Pan, B.; Xia, Y.; Liu, L. Network pharmacology-based analysis reveals the putative action mechanism of Polygonum cuspidatum against COVID-19. Int. J. Clin. Exp. Med. 2021, 14, 1852–1863. [Google Scholar]
- Xu, H.; Li, J.; Song, S.; Xiao, Z.; Chen, X.; Huang, B.; Sun, M.; Su, G.; Zhou, D.; Wang, G.; et al. Effective inhibition of coronavirus replication by Polygonum cuspidatum. Front. Biosci. 2021, 26, 789–798. [Google Scholar] [CrossRef]
- Yu, M.X.; Song, X.; Ma, X.Q.; Hao, C.X.; Huang, J.J.; Yang, W.H. Investigation into molecular mechanisms and high-frequency core TCM for pulmonary fibrosis secondary to COVID-19 based on network pharmacology and data mining. Ann. Palliat. Med. 2021, 10, 3960–3975. [Google Scholar] [CrossRef]
- Wang, M.; Qin, K.; Zhai, X. Combined network pharmacology, molecular docking, and experimental verification approach to investigate the potential mechanisms of polydatin against COVID-19. Nat. Prod. Commun. 2022, 17, 1934578X221095352. [Google Scholar] [CrossRef]
- Pasquereau, S.; Nehme, Z.; Haidar Ahmad, S.; Daouad, F.; Van Assche, J.; Wallet, C.; Schwartz, C.; Rohr, O.; Morot-Bizot, S.; Herbein, G. Resveratrol inhibits HCoV-229E and SARS-CoV-2 Coronavirus replication in vitro. Viruses 2021, 13, 354. [Google Scholar] [CrossRef]
- Wiwanitkit, V. Polydatin and COVID-19. Clin. Nutr. [CrossRef]
- Naumoska, K.; Jug, U.; Kõrge, K.; Oberlintner, A.; Golob, M.; Novak, U.; Vovk, I.; Likozar, B. Antioxidant and antimicrobial biofoil based on chitosan and Japanese knotweed (Fallopia japonica, Houtt.) rhizome bark extract. Antioxidants 2022, 11, 1200. [Google Scholar] [CrossRef]
- Van Doremalen, N.; Bushmaker, T.; Morris, D.H.; Holbrook, M.G.; Gamble, A.; Williamson, B.N.; Tamin, A.; Harcourt, J.L.; Thornburg, N.J.; Gerber, S.I.; et al. Aerosol and surface stability of SARS-CoV-2 as compared with SARS-CoV-1. N. Engl. J. Med. 2020, 382, 1564–1567. [Google Scholar] [CrossRef] [PubMed]
- Why the Coronavirus Has Been So Successful. Available online: https://www.theatlantic.com/science/archive/2020/03/biography-new-coronavirus/608338/ (accessed on 14 November 2021).
- Meyers, C.; Kass, R.; Goldenberg, D.; Milici, J.; Alam, S.; Robison, R. Ethanol and isopropanol inactivation of human coronavirus on hard surfaces. J. Hosp. Infect. 2021, 107, 45–49. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Graph showing the reduction in CPE by 70% ethanol(aq) Japanese knotweed rhizome bark extract on SARS-CoV-2 performed in 48 parallels at extract concentrations that did not cause cytotoxicity in comparison with the optimal working concentration of the virus (6.2 TCID50 SARS-CoV-2). The results are presented as a percentage of wells with cytopathic effect on Vero-E6 cells (n = 48); * p < 0.05. The result for the positive control (cells exposed to the virus only) is presented for comparison.
Figure 1.
Graph showing the reduction in CPE by 70% ethanol(aq) Japanese knotweed rhizome bark extract on SARS-CoV-2 performed in 48 parallels at extract concentrations that did not cause cytotoxicity in comparison with the optimal working concentration of the virus (6.2 TCID50 SARS-CoV-2). The results are presented as a percentage of wells with cytopathic effect on Vero-E6 cells (n = 48); * p < 0.05. The result for the positive control (cells exposed to the virus only) is presented for comparison.

Figure 2.
Images of: (A) non-treated Vero-E6 cells; (B) Vero-E6 cells exposed to SARS-CoV-2 virus (6.2 TCID50 SARS-CoV-2) and Japanese knotweed rhizome bark extract in concentration which prevents a cytopathic effect without provoking cytotoxicity (50.8 µg mL−1); (C) cytotoxic effect caused by higher concentrations of Japanese knotweed rhizome bark extract; (D) cytopathic effect resulting from viral infection.
Figure 2.
Images of: (A) non-treated Vero-E6 cells; (B) Vero-E6 cells exposed to SARS-CoV-2 virus (6.2 TCID50 SARS-CoV-2) and Japanese knotweed rhizome bark extract in concentration which prevents a cytopathic effect without provoking cytotoxicity (50.8 µg mL−1); (C) cytotoxic effect caused by higher concentrations of Japanese knotweed rhizome bark extract; (D) cytopathic effect resulting from viral infection.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jug, U.; Naumoska, K.; Malovrh, T. Japanese Knotweed Rhizome Bark Extract Inhibits Live SARS-CoV-2 In Vitro. Bioengineering 2022, 9, 429. https://doi.org/10.3390/bioengineering9090429
AMA Style
Jug U, Naumoska K, Malovrh T. Japanese Knotweed Rhizome Bark Extract Inhibits Live SARS-CoV-2 In Vitro. Bioengineering. 2022; 9(9):429. https://doi.org/10.3390/bioengineering9090429
Chicago/Turabian StyleJug, Urška, Katerina Naumoska, and Tadej Malovrh. 2022. "Japanese Knotweed Rhizome Bark Extract Inhibits Live SARS-CoV-2 In Vitro" Bioengineering 9, no. 9: 429. https://doi.org/10.3390/bioengineering9090429
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.