
Buckwheat Milling Waste Effects on Root Morphology and Mycorrhization of Silver Fir Seedlings Inoculated with Black Summer Truffle (Tuber aestivum Vittad.)

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
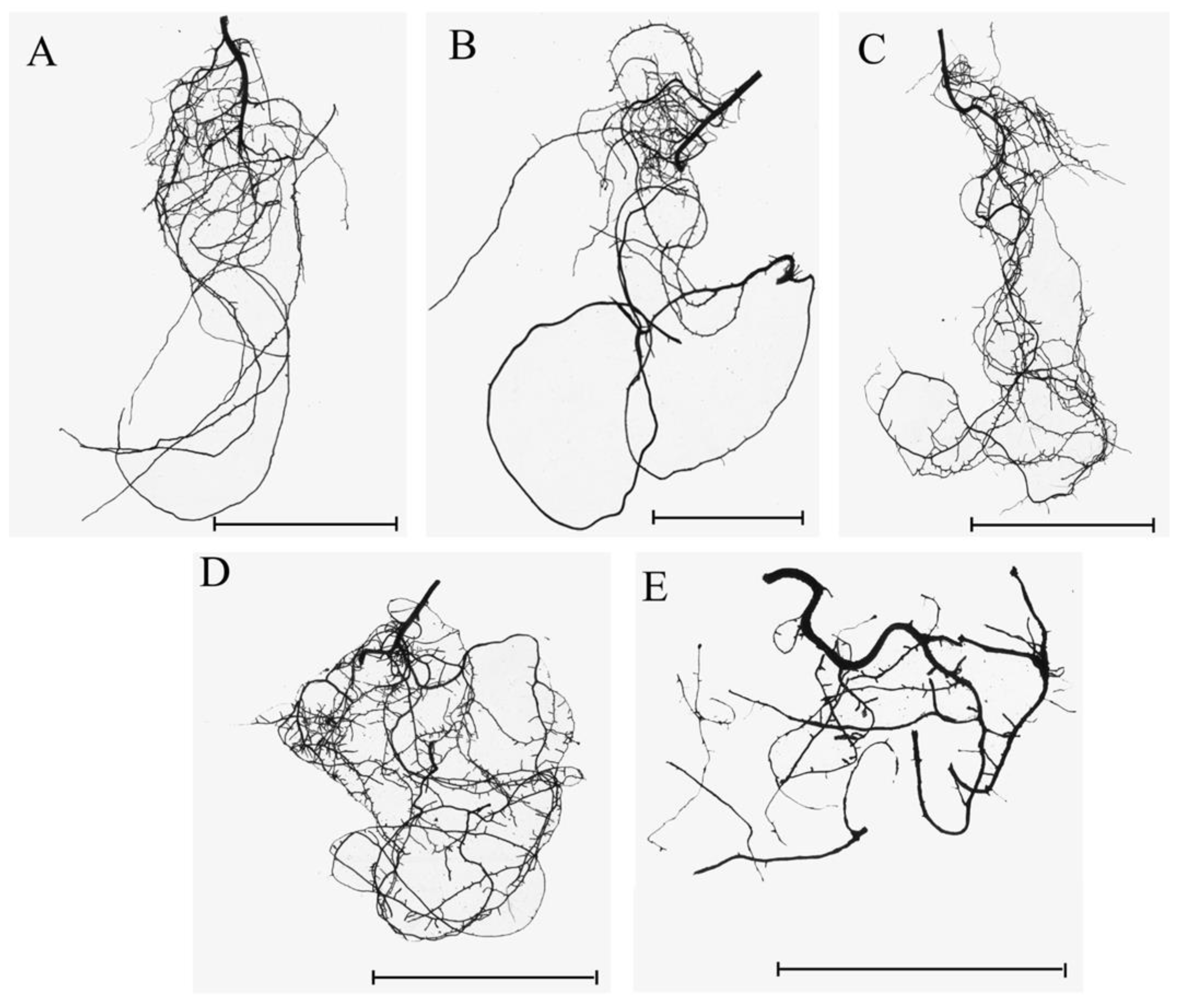
2.2. Root Morphology Assessment and Evaluation of Seedling Mycorrhization Level
2.3. Molecular Analysis
2.4. Basic Chemical Analyses of Substrate
2.5. Rutin and Quercetin Concentration in Buckwheat Husks/Bran
2.6. Statistical Analyses
3. Results and Discussion
3.1. Chemical Composition of the Substrate
3.2. Rutin and Quercetin Concentrations in Buckwheat By-Products
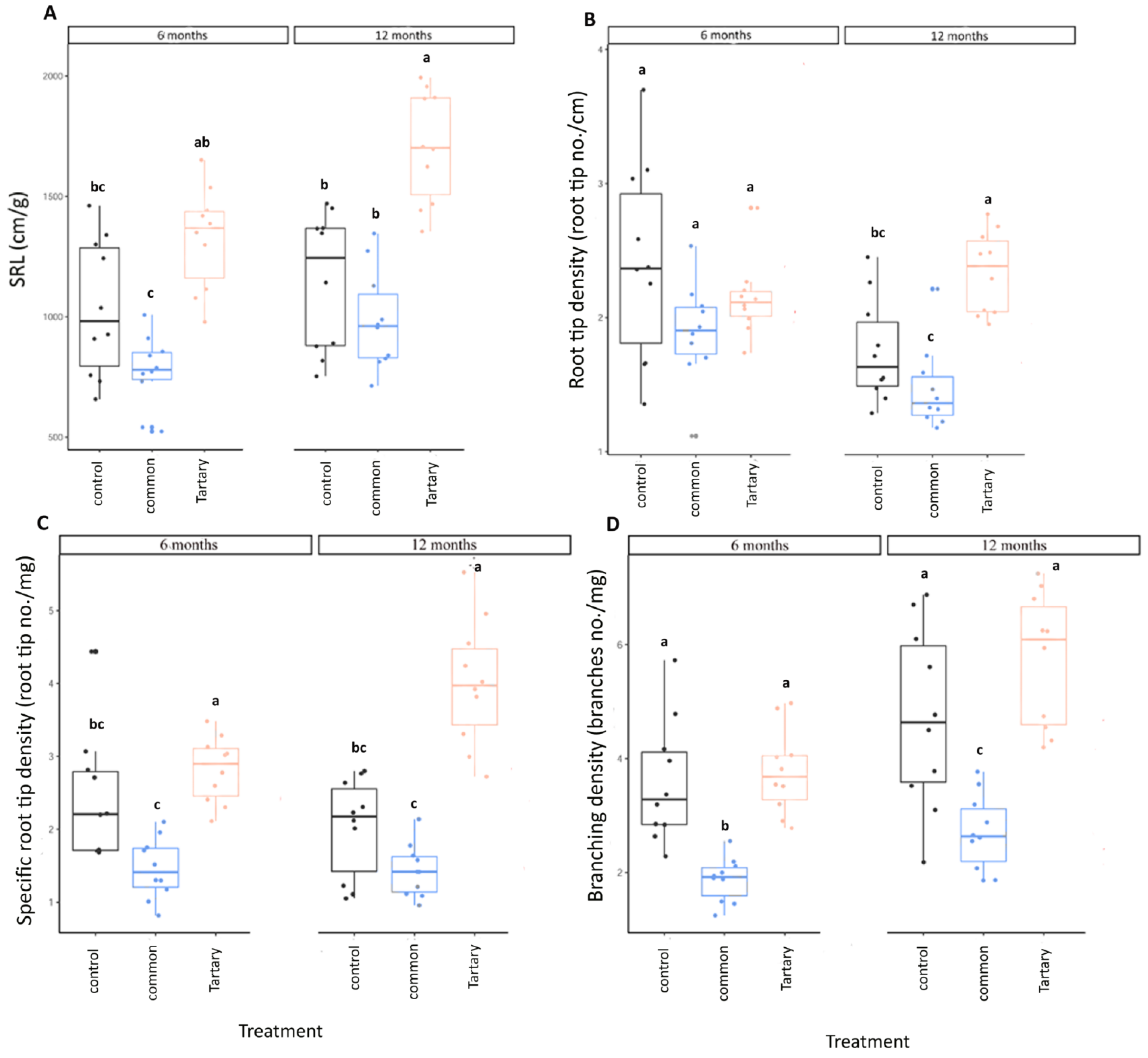
3.3. Impact of Husks on Root Growth
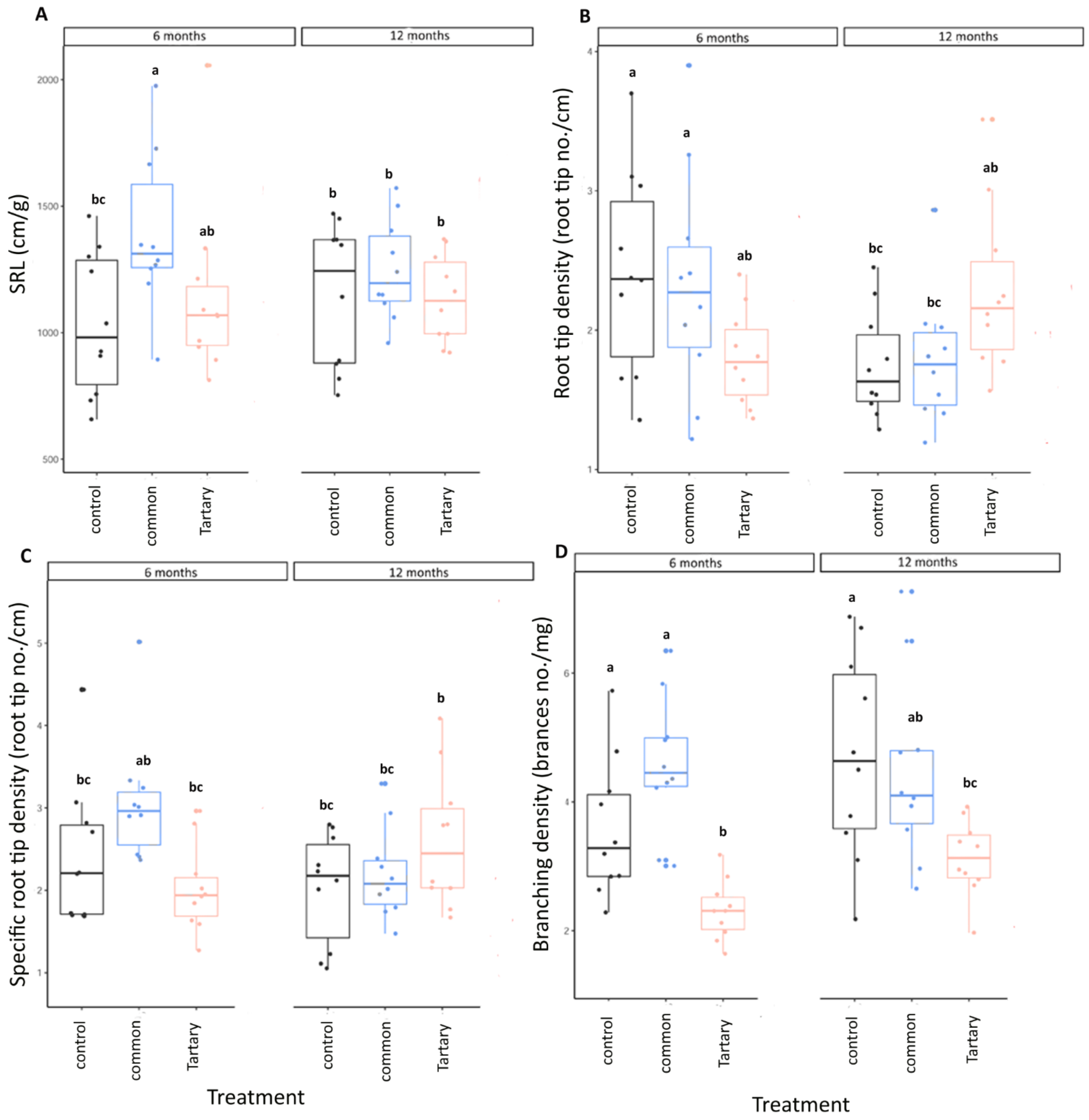
3.4. Impact of Bran on Root Growth
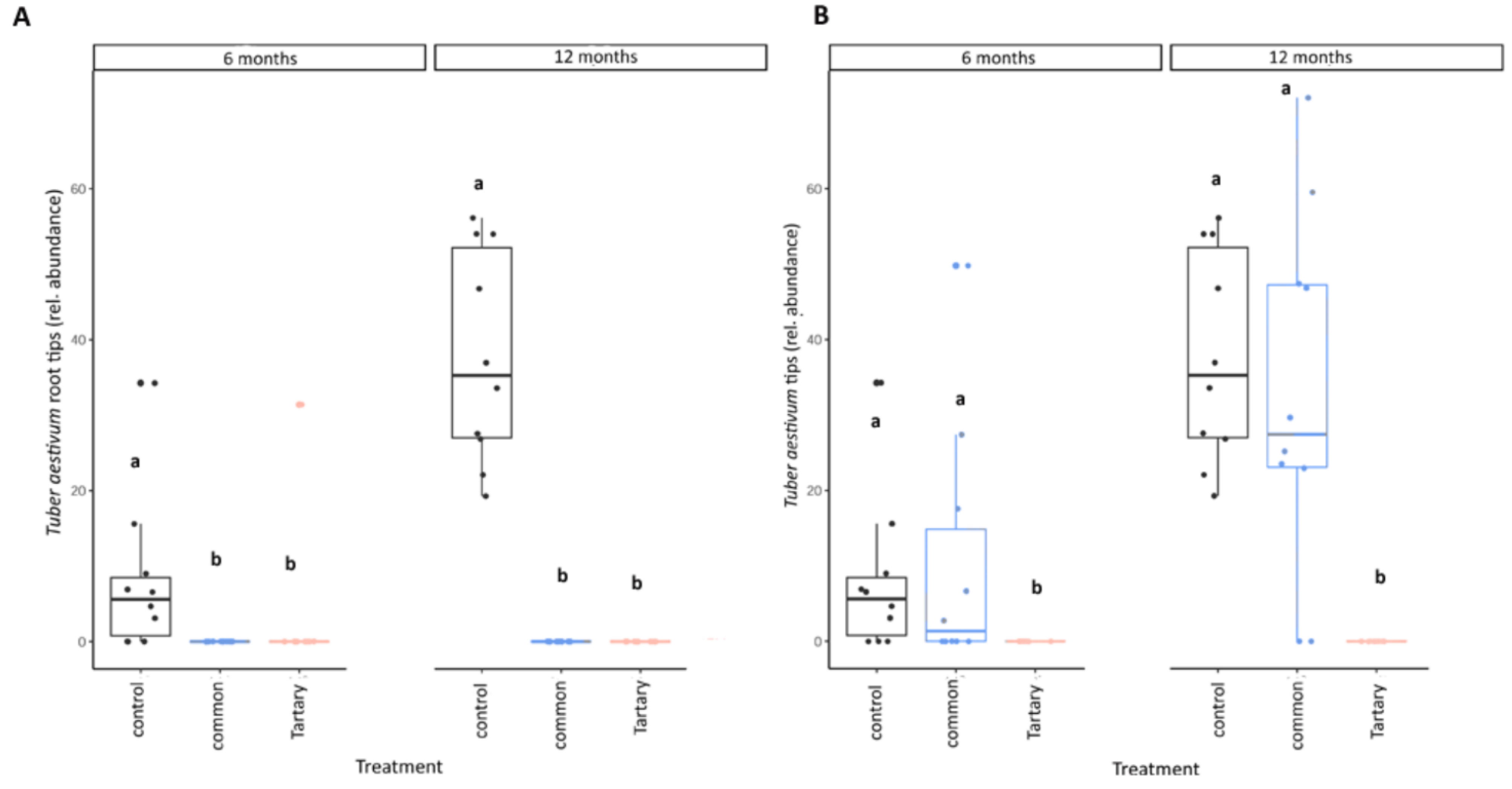
3.5. Buckwheat Husks Had a Negative Impact on Summer Truffle Mycorrhization Level
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. The Food and Agriculture Organization of the United Nation. Available online: http://www.fao.org/faostat/en/#data (accessed on 15 December 2021).
- Dziedzic, K.; Górecka, D.; Szwengiel, A.; Olejnik, A.; Rychlik, J.; Kreft, I.; Drożdżyńska, A.; Walkowiak, J. The cytotoxic effect of artificially digested buckwheat products on HT-29 colon cancer cells. J. Cereal Sci. 2018, 83, 68–73. [Google Scholar] [CrossRef]
- Dziedzic, K.; Górecka, D.; Szwengiel, A.; Sulewska, H.; Kreft, I.; Gujska, E.; Walkowiak, J. The Content of Dietary Fibre and Polyphenols in Morphological Parts of Buckwheat (Fagopyrum tataricum). Plant Foods Hum. Nutr. 2018, 73, 82–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SiStat Production of Crops (ha, T, t/ha), Slovenia, Annually. Available online: https://pxweb.stat.si/SiStatDb/pxweb/en/30_Okolje/30_Okolje__15_kmetijstvo_ribistvo__04_rastlinska_pridelava__01_15024_pridelki_povrsina/1502402S.px/ (accessed on 18 December 2021).
- Bonafaccia, G.; Marocchini, M.; Kreft, I. Composition and technological properties of the flour and bran from common and tartary buckwheat. Food Chem. 2003, 80, 9–15. [Google Scholar] [CrossRef]
- Faulds, C.B.; Bartolomé, B.; Williamson, G. Novel biotransformations of agro-industrial cereal waste by ferulic acid esterases. Ind. Crops Prod. 1997, 6, 367–374. [Google Scholar] [CrossRef]
- Ramirez, E.C.; Johnston, D.B.; McAloon, A.J.; Yee, W.; Singh, V. Engineering process and cost model for a conventional corn wet milling facility. Ind. Crops Prod. 2008, 27, 91–97. [Google Scholar] [CrossRef]
- Theerarattananoon, K.; Xu, F.; Wilson, J.; Ballard, R.; Mckinney, L.; Staggenborg, S.; Vadlani, P.; Pei, Z.J.; Wang, D. Physical properties of pellets made from sorghum stalk, corn stover, wheat straw, and big bluestem. Ind. Crops Prod. 2011, 33, 325–332. [Google Scholar] [CrossRef]
- Teh, C.Y.; Wu, T.Y.; Juan, J.C. Optimization of agro-industrial wastewater treatment using unmodified rice starch as a natural coagulant. Ind. Crops Prod. 2014, 56, 17–26. [Google Scholar] [CrossRef]
- Viscusi, G.; Adami, R.; Gorrasi, G. Fabrication of rice flour films reinforced with hemp hurd and loaded with grapefruit seed oil: A simple way to valorize agro-waste resources toward low cost materials with added value. Ind. Crops Prod. 2021, 170, 113785. [Google Scholar] [CrossRef]
- Kreft, I.; Zhou, M.; Golob, A.; Germ, M.; Likar, M.; Dziedzic, K.; Luthar, Z. Breeding buckwheat for nutritional quality. Breed. Sci. 2020, 70, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Kreft, I.; Chao, G.; Wang, Y.; Liu, X.; Wang, L.; Wang, P.; Gao, X.; Feng, B. Tartary buckwheat (Fagopyrum tataricum Gaertn.) starch, a side product in functional food production, as a potential source of retrograded starch. Food Chem. 2016, 190, 552–558. [Google Scholar] [CrossRef]
- Kreft, M. Buckwheat phenolic metabolites in health and disease. Nutr. Res. Rev. 2016, 29, 30–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukšič, L.; Bonafaccia, G.; Timoracka, M.; Vollmannova, A.; Trček, J.; Nyambe, T.K.; Melini, V.; Acquistucci, R.; Germ, M.; Kreft, I. Rutin and quercetin transformation during preparation of buckwheat sourdough bread. J. Cereal Sci. 2016, 69, 71–76. [Google Scholar] [CrossRef]
- Ikeda, K.; Oku, M.; Kusano, T.; Yasumoto, K. Inhibitory Potency of Plant Antinutrients towards the In Vitro Digestibility of Buckwheat Protein. J. Food Sci. 1986, 51, 1527–1530. [Google Scholar] [CrossRef]
- Skrabanja, V.; Lærke, H.N.; Kreft, I. Protein-polyphenol interactions and in vivo digestibility of buckwheat groat proteins. Pflügers Arch.-Eur. J. Physiol. 2000, 440, R129–R131. [Google Scholar] [CrossRef] [PubMed]
- Nahberger, T.U.; Benucci, G.M.N.; Kraigher, H.; Grebenc, T. Effect of earthworms on mycorrhization, root morphology and biomass of silver fir seedlings inoculated with black summer truffle (Tuber aestivum Vittad.). Sci. Rep. 2021, 11, 6167. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.; Colinas, C. Methology for certification of Quercus ilex seedlings inoculated with Tuber melanosporum for commercial application. In Proceedings of the First International Conference on Mycorrhiza, Berkeley CA, USA, 4–9 August 1996. [Google Scholar]
- Unuk Nahberger, T.; Kraigher, H.; Grebenc, T. PCR primers comparisons for a successful Tuber spp. DNA region amplification in routine identifications/Primerjava PCR začetnih oligonukleotidov za uspešno pomnoževanje DNA regije Tuber spp. pri rutinski identifikaciji. Folia Biol. Geol. 2020, 61, 229–238. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal rna genes for phylogenetics. In PCR Protocols; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Bertini, L. A new pair of primers designed for amplification of the ITS region in Tuber species. FEMS Microbiol. Lett. 1999, 173, 239–245. [Google Scholar] [CrossRef] [Green Version]
- Grebenc, T.; Kraigher, H. Types of ectomycorrhiza of mature beech and spruce at ozone-fumigated and control forest plots. Environ. Monit. Assess. 2007, 128, 47–59. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2016. [Google Scholar]
- Cho, Y.J.; Bae, I.Y.; Inglett, G.E.; Lee, S. Utilization of tartary buckwheat bran as a source of rutin and its effect on the rheological and antioxidant properties of wheat-based products. Ind. Crops Prod. 2014, 61, 211–216. [Google Scholar] [CrossRef]
- Vojtíšková, P.; Švec, P.; Kubáň, V.; Krejzová, E.; Bittová, M.; Kráčmar, S.; Svobodová, B. Chemical composition of buckwheat plant parts and selected buckwheat products. Potravinarstvo 2014, 8, 247–253. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-J.; Zaidul, I.S.M.; Suzuki, T.; Mukasa, Y.; Hashimoto, N.; Takigawa, S.; Noda, T.; Matsuura-Endo, C.; Yamauchi, H. Comparison of phenolic compositions between common and tartary buckwheat (Fagopyrum) sprouts. Food Chem. 2008, 110, 814–820. [Google Scholar] [CrossRef]
- Fabjan, N.; Rode, J.; Košir, I.J.; Wang, Z.; Zhang, Z.; Kreft, I. Tartary Buckwheat (Fagopyrum tataricum Gaertn.) as a Source of Dietary Rutin and Quercitrin. J. Agric. Food Chem. 2003, 51, 6452–6455. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, V.P.; Chauhan, D.K.; Prasad, S.M.; Dubey, N.K. Improvement of Crops in the Era of Climatic Changes; Ahmad, P., Wani, M.R., Azooz, M.M., Phan Tran, L.-S., Eds.; Springer: New York, NY, USA, 2014; ISBN 978-1-4614-8823-1. [Google Scholar]
- Hirsch, A.M.; Fang, Y.; Asad, S.; Kapulnik, Y. The role of phytohormones in plant-microbe symbioses. Plant Soil 1997, 194, 171–184. [Google Scholar] [CrossRef]
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Viégas, I.d.J.M.; Naif, A.P.M.; da Conceição, H.E.O.; Lobato, A.K.d.S.; Frazão, D.A.C.; Neto, C.F.d.O.; Cordeiro, R.A.M. Visual symptoms, growth and nutrients of Alpinia purpurata plants exposed to N, P, K, Ca, Mg and S deficiencies. J. Food Agric. Environ. 2011, 9, 1048–1051. [Google Scholar]
- Brown, D.E.; Rashotte, A.M.; Murphy, A.S.; Normanly, J.; Tague, B.W.; Peer, W.A.; Taiz, L.; Muday, G.K. Flavonoids Act as Negative Regulators of Auxin Transport in Vivo in Arabidopsis. Plant Physiol. 2001, 126, 524–535. [Google Scholar] [CrossRef] [Green Version]
- Murphy, A.; Peer, W.A.; Taiz, L. Regulation of auxin transport by aminopeptidases and endogenous flavonoids. Planta 2000, 211, 315–324. [Google Scholar] [CrossRef]
- Peer, W.A.; Murphy, A.S. Flavonoids as Signal Molecules: Targets of Flavonoid Action. In The Science of Flavonoids; Springer: New York, NY, USA, 2006; pp. 239–268. ISBN 9780387288222. [Google Scholar]
- Peer, W.A.; Murphy, A.S. Flavonoids and auxin transport: Modulators or regulators? Trends Plant Sci. 2007, 12, 556–563. [Google Scholar] [CrossRef]
- Teale, W.; Palme, K. Naphthylphthalamic acid and the mechanism of polar auxin transport. J. Exp. Bot. 2018, 69, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Lomax, T.L.; Muday, G.K.; Rubery, P.H. Auxin Transport. In Plant Hormones; Davies, P.J., Ed.; Springer: Dordrecht, The Netherlands, 1995; pp. 509–530. [Google Scholar]
- Chevalier, G.; Sourzat, P. Soils and Techniques for Cultivating Tuber melanosporum and Tuber aestivum in Europe. In Edible Ectomycorrhizal Mushrooms; Zambonelli, A., Bonito, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 163–189. [Google Scholar]
- Fischer, C.; Oliach, D.; Bonet, J.A.; Colinas, C. Best Practices for Cultivation of Truffles; Centre Tecnològic Forestal de Catalunya: Solsona, Spain, 2017. [Google Scholar]
- García-Montero, L.G.; Casermeiro, M.A.; Hernando, I.; Hernando, J. Effect of active carbonate, exchangeable calcium, and stoniness of soil on Tuber melanosporum carpophore production. N. Zeal. J. Crop Hortic. Sci. 2007, 35, 139–146. [Google Scholar] [CrossRef]
- Li, F.H.; Yuan, Y.; Yang, X.L.; Tao, S.Y.; Ming, J. Phenolic Profiles and Antioxidant Activity of Buckwheat (Fagopyrum esculentum Möench and Fagopyrum tartaricum L. Gaerth) Hulls, Brans and Flours. J. Integr. Agric. 2013, 12, 1684–1693. [Google Scholar] [CrossRef] [Green Version]
- Čabarkapa, I.S.; Sedej, I.J.; Sakač, M.B.; Šarić, L.Ć.; Plavšič, D.V. Antimicrobial activity of buckwheat. Food Process. Qual. Saf. 2008, 35, 159–163. [Google Scholar]
- Wang, L.; Yang, X.; Qin, P.; Shan, F.; Ren, G. Flavonoid composition, antibacterial and antioxidant properties of tartary buckwheat bran extract. Ind. Crops Prod. 2013, 49, 312–317. [Google Scholar] [CrossRef]
- Veras, H.N.H.; Dos Santos, I.J.M.; Dos Santos, A.C.B.; Fernandes, C.N.; Marias, E.F.F.; Leite, G.D.O.; De Souza, H.H.F.; Da Costa, J.G.M.; Coutinho, H.D.M. Comparative evaluation of antibiotic and antibiotic modifying activity of quercetin and isoquercetin in vitro. Curr. Top. Nutraceutical Res. 2011, 9, 25–29. [Google Scholar]
- Hattori, M.; Yamaji-Tsukamoto, K.; Kumagai, H.; Feng, Y.; Takahashi, K. Antioxidative Activity of Soluble Elastin Peptides. J. Agric. Food Chem. 1998, 46, 2167–2170. [Google Scholar] [CrossRef]
- Cushnie, T.P.T.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef]
- Zabka, M.; Pavela, R. Antifungal efficacy of some natural phenolic compounds against significant pathogenic and toxinogenic filamentous fungi. Chemosphere 2013, 93, 1051–1056. [Google Scholar] [CrossRef]
- Ponts, N.; Pinson-Gadais, L.; Boutigny, A.L.; Barreau, C.; Richard-Forget, F. Cinnamic-derived acids significantly affect fusarium graminearum growth and in vitro synthesis of type B trichothecenes. Phytopathology 2011, 101, 929–934. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, L.; Bonnin-Verdal, M.N.; Marchegay, G.; Pinson-Gadais, L.; Ducos, C.; Richard-Forget, F.; Atanasova-Penichon, V. Fungal biotransformation of chlorogenic and caffeic acids by Fusarium graminearum: New insights in the contribution of phenolic acids to resistance to deoxynivalenol accumulation in cereals. Int. J. Food Microbiol. 2016, 221, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Kumaraswamy, G.K.; Bollina, V.; Kushalappa, A.C.; Choo, T.M.; Dion, Y.; Rioux, S.; Mamer, O.; Faubert, D. Metabolomics technology to phenotype resistance in barley against Gibberella zeae. Eur. J. Plant Pathol. 2011, 130, 29–43. [Google Scholar] [CrossRef]
- Schöneberg, T.; Kibler, K.; Sulyok, M.; Musa, T.; Bucheli, T.D.; Mascher, F.; Bertossa, M.; Voegele, R.T.; Vogelgsang, S. Can plant phenolic compounds reduce Fusarium growth and mycotoxin production in cereals? Food Addit. Contam.-Part A Chem. Anal. Control. Expo. Risk Assess. 2018, 35, 2455–2470. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time after Spore Inoculation (Months) | Treatment | Organic Carbon Content (%) | Mineral Carbon Content (%) | Nitrogen Content (%) | C/N Ratio |
|---|---|---|---|---|---|
| zero point | Control | 22.5 ± 0.5a | 1.4 ± 0.01c | 0.505 ± 0.006b | 44:1 |
| zero point | CB bran | 23.6 ± 0.4a | 1.8 ± 0.02b | 0.817 ± 0.008a | 29:1 |
| zero point | TB bran | 17.2 ± 0.5bc | 2.6 ±0.05a | 0.980 ± 0.03a | 18:1 |
| zero point | CB husks | 19.4 ± 0.07b | 1.9 ± 0.03b | 0.426 ± 0.001bc | 45:1 |
| zero point | TB husks | 15.6 ± 0.3c | 2 ± 0.01b | 0.354 ± 0.008c | 44:1 |
| 6 | Control | 19.5 ± 0.1b | 1.2 ± 0.03b | 0.455 ± 0.002b | 43:1 |
| 6 | CB bran | 21.5 ± 0.2a | 1.6 ± 0.02b | 1.027 ± 0a | 21:1 |
| 6 | TB bran | 14.7 ± 0.1c | 2.2 ± 0.1a | 0.888 ± 0.01a | 17:1 |
| 6 | CB husks | 19.7 ± 0.2b | 1.5 ± 0.02b | 0.496 ± 0.009b | 40:1 |
| 6 | TB husks | 14.8 ± 0.1c | 2.1 ± 0.09a | 0.400 ± 0.08b | 37:1 |
| 12 | Control | 17.7 ± 0.3b | 1.4 ± 0.02b | 0.418 ± 0.009b | 42:1 |
| 12 | CB bran | 21.6 ± 0.03a | 1.5 ± 0.2b | 0.770 ± 0.01a | 28:1 |
| 12 | TB bran | 12.6 ± 0.02c | 2.0 ± 0.07ab | 0.679 ± 0.008a | 19:1 |
| 12 | CB husks | 18.4 ± 0.2b | 1.6 ± 0.01b | 0.501 ± 0.004ab | 37:1 |
| 12 | TB husks | 13.3 ± 0.2c | 2.2 ± 0.1a | 0.382 ± 0.01b | 35:1 |
| Buckwheat Product | Rutin (mg/g Dry Weight) | Quercetin (mg/g Dry Weight) |
|---|---|---|
| CB bran | 0.08 ± 0.04b | 0.012 ± 0.002b |
| TB bran | 0.51 ± 0.02a | 0.701 ± 0.021a |
| CB husks | 0.02 ± 0.03b | 0.010 ± 0.006b |
| TB husks | 0.07 ± 0.02b | 0.031 ± 0.001b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nahberger, T.U.; Grebenc, T.; Žlindra, D.; Mrak, T.; Likar, M.; Kraigher, H.; Luthar, Z. Buckwheat Milling Waste Effects on Root Morphology and Mycorrhization of Silver Fir Seedlings Inoculated with Black Summer Truffle (Tuber aestivum Vittad.). Forests 2022, 13, 240. https://doi.org/10.3390/f13020240
Nahberger TU, Grebenc T, Žlindra D, Mrak T, Likar M, Kraigher H, Luthar Z. Buckwheat Milling Waste Effects on Root Morphology and Mycorrhization of Silver Fir Seedlings Inoculated with Black Summer Truffle (Tuber aestivum Vittad.). Forests. 2022; 13(2):240. https://doi.org/10.3390/f13020240
Chicago/Turabian StyleNahberger, Tina Unuk, Tine Grebenc, Daniel Žlindra, Tanja Mrak, Matevž Likar, Hojka Kraigher, and Zlata Luthar. 2022. "Buckwheat Milling Waste Effects on Root Morphology and Mycorrhization of Silver Fir Seedlings Inoculated with Black Summer Truffle (Tuber aestivum Vittad.)" Forests 13, no. 2: 240. https://doi.org/10.3390/f13020240