
Potential Link between Ectomycorrhizal Community Composition and Host Tree Phenology


Department of Forest Physiology and Genetics, Slovenian Forestry Institute, 1000 Ljubljana, Slovenia
*
Author to whom correspondence should be addressed.
Forests 2021, 12(12), 1719; https://doi.org/10.3390/f12121719
Submission received: 30 June 2021
/
Revised: 29 November 2021
/
Accepted: 4 December 2021
/
Published: 7 December 2021
(This article belongs to the Special Issue Forests, Their Diversity, and Conservation)
Abstract
:The timing of leaf phenology tends to be crucial in controlling ecosystem processes such as the acquisition of carbon and water loss as well as in controlling tree nutrient cycling. To date, tree phenology has mostly been associated with environmental control (e.g., temperature and photoperiod) in a relationship with inheritance, while it has rarely been linked with ectomycorrhizal community compositional changes through the host tree’s phenological stages. Seasonal variations of fungal communities have been widely studied, but little is known about mycorrhiza community composition changes along phenological stages. Therefore, we analyzed ectomycorrhizal communities associated with silver fir and their compositional changes during the transition between phenological stages. The phenological stages of each individual tree and time of bud break were monitored weekly for two years and, at the same time, ectomycorrhiza was harvested from selected silver fir trees. In total, 60 soil cores were analyzed for differences in the ectomycorrhizal community between phenological stages using Sanger sequencing of individual ectomycorrhizal morphotypes. A significant difference in beta diversity for an overall ectomycorrhizal community was confirmed between analyzed time periods for both sampled years. Species-specific reactions to transitions between phenological stages were confirmed for 18 different ectomycorrhizal taxa, where a positive correlation of Russula ochroleuca, Russula illota, Tomentella sublilacina, and Tylospora fibrillosa was observed with the phenological stage of bud burst.
1. Introduction
Silver fir (Abies alba Mill.) is one of the most valuable relic conifer tree species in Europe [1]. It is considered a significant ecological and functional balancer tree species of European forests. Among other functions, it stabilizes soils and retains water [2]. However, little is known about its ectomycorrhizal symbionts and their ecological and physiological role in providing vigor to silver fir [1]. Furthermore, to date, ectomycorrhizal community shifts have rarely been linked to tree leaf phenology, with some of these studies only focusing on deciduous trees. Leaf phenology is an important trait affecting tree survival and growth through its impact on carbon balance and overall tree fitness [3,4]. Due to environmental conditions and endogenous rhythms, tree phenology changes seasonally, from bud bursting to full leaf/needle expansion over a growing season followed by a period of winter dormancy [5]. To date, the timing of belowground organs’ phenology, such as fine roots, paralleled to leaf phenology has received less attention even though it likely influences overall plant functioning and fitness [6].
Among others, mycorrhizal symbiosis is an important interaction and should not be ignored in phenological studies. For instance, root colonization by ectomycorrhiza has been shown to correlate with earlier bud burst in oak seedlings [7] and in our previous study we confirmed a significant difference in ectomycorrhizal community composition between early bud bursting and late bud bursting silver fir seedlings [8].
Although tree phenology and seasonal variations of fungal communities have been widely studied, their interdependence—the above–below ground link between tree phenology and ectomycorrhizal fungal community composition shifts—is largely unknown. Only scarce, early studies have correlated ectomycorrhizal root colonization and tree phenology [7,9,10]. Ectomycorrhizalroot colonization most likely affects host phenology through improved mineral absorption capacity of mycorrhizal roots; Courty et al. [11] and Waldrop et al. [12] strongly suggested a mycorrhizal effect on host phenology by providing the carbon to the host tree at a time when demand is high and photoassimilates are not yet available. At the time of bud break, when carbon supply is provided exclusively by the metabolic activity of stored reserves, Dhuli et al. [13] and Courty et al. [11] recorded an increase in organic matter degrading enzymes produced by Lactarius quietus just before bud break, which mobilize carbon as glucose at the time of formation of new leaves. Changes in mycorrhizal communities throughout the season also affect nutrient and water availability, and thus directly impact the host tree [14].
Short-term shifts in the ectomycorrhizal community in relation to the ectomycorrhizal tree partner phenology stage of development had not yet been analyzed on a detailed time scale, thus we analyzed ectomycorrhizal fungal community composition changes along the transitions between phenological stages and species-specific reactions to those transitions. We described the ectomycorrhizal fungal community per individual phenological stage of silver fir in native silver fir-dominated forest and hypothesized that the whole ectomycorrhizal community structure will differ between phenological stages. At least for some in our study-identified ectomycorrhizal taxa, species-specific reactions to transitions between silver fir phenological stages are expected.
2. Materials and Methods
2.1. Study Site and Phenological Observations
This study was performed in a silver fir (Abies alba Mill.)-dominated forest site measuring 16.47 hectares and located in the ‘Lehen na Pohorju’ area (Pohorje, Slovenia, 46°33′ N 15°20′ E, 469–611 m a. s. l.). The phenological stages of silver fir trees in natural regenerating centers were assessed based on a simplified protocol [15] by visual observations for two years (2016 and 2017). Phenological observations of bud break were carried out once per week. Phenological stages were simplified and determined as (1) dormancy (buds enclosed by needles and not visible), (2) bud bursting (bud elongation, and elongating brush of soft needles emerge), and (3) full needle expansion (shoots with developed needles). Soil/root cores for ectomycorrhizal identification were collected at three different time periods, which were linked to host tree phenology (Table 1).
The observations of phenological stages ended when the final phenological stage was confirmed for all analyzed silver fir trees.
2.2. Mycorrhizal Sampling
Ectomycorrhiza sampling collection times were determined by the phenological stage of individual silver fir trees. Ectomycorrhizal samples were harvested by sequential soil coring with a soil corer from ten preselected mature silver fir trees. Each soil core was 3.2 cm in diameter and 20 cm deep. The sampling methodology followed that of Kraigher [16]. Altogether, we collected 60 soil samples in three different time periods and in 2 consecutive years (Table 1). To avoid roots sampling from other adult tree species, soil samples were collected approximately 1 m from the trunk, whereas individual silver fir trees were at least 5–7 m apart to minimize sampling from the same common mycorrhizal network [17,18,19,20].
Individual soil core samples were placed in plastic bags and stored at 4 °C before being processed. All roots from individual soil core samples were soaked in tap water for approximately 15–20 min before being gently washed to remove all soil particles attached to the roots. For further processing, the roots from individual soil core samples were divided into coarse or fine roots. Fine roots were defined as roots with a diameter of less than 2 mm [19,21]. Fine roots were observed in tap water under an Olympus SZX12 stereomicroscope (Olympus, Hamburg, Germany). Altogether, sampling resulted in 105.085 ectomycorrhizal root tips characterized and included in further analyses. The pieces of fine roots were placed above a transparent plexi-glass tray with a grid. The roots were equally distributed throughout the grid, from which one further square was randomly chosen. Within the selected square, all root tips were characterized, counted, and separated into vital and non-vital (dead) ectomycorrhizal root tips. The number of root tips counted and characterized in each sample was 500. Vital ectomycorrhizal root tips were further separated based on morphological and anatomical features and characterized according to the method of Agerer [22]. Although non-vital ectomycorrhizal root tips could be due to specific morpho-anatomical characteristics categorized into morphotype, we ranked them among non-vital/dead ectomycorrhizal root tips. Altogether, we characterized 30,000 ectomycorrhizal root tips, including vital and non-vital (dead) mycorrhizas. The relative abundance of individual ectomycorrhizal morphotypes was thereby calculated as the number of individual ectomycorrhizal morphotype root tips/500 analyzed root tips. Further, three to five ectomycorrhizal root tips of each morphotype from individual soil core samples were kept at −20 °C and later freeze-dried at −92 °C for 48 h. After freeze-drying, roots were homogenized into a fine powder using a Retsch mixer mill (Retsch, Hahn, Germany) and analyzed using molecular methods.
2.3. Molecular Analysis
Total DNA from each morphotype was extracted with a DNeasy Plant Mini kit (Qiagen, Hilden, Germany), following the manufacturer’s instructions. The complete ITS region of nuclear ribosomal DNA was amplified from isolated DNA using the fungus specific primer pair ITS1F and ITS4 [23,24], following the modified procedure described in Sulzbacher et al. [25]. PCR products were run on 1.5% agarose gels in 0.5× TBE buffer and visualized with Gel Doc EQ System, PC (BioRad, Hercules, CA, USA). For morphotypes for which amplification failed, the extracted DNA was diluted 10:1 and re-amplified. The amplification success of morphotypes varies between 78% and 100% per individual sample. Amplified DNA fragments were cut from an agarose gel and purified with innuPREP DOUBLEpure Kit (Analytik Jena AG, Jena, Germany), following the manufacturer’s instructions. After amplified DNA fragment purification, Sangar sequencing was performed at a commercial sequencing laboratory (Macrogen Inc., Seoul, Korea). All morphotypes were sequenced in both directions with ITS1F and ITS4 primers [23,24]. The obtained sequences were processed in Geneious version 11.1.4 (https://www.geneious.com, accessed on 3 September 2019) [26]. Nucleotide base calls with an error probability greater than 5% were trimmed from read ends to improve read quality, while reads were assembled into contigs at 90% base pair similarity. The BLASTN algorithm from the NCBI website (National Center for Biotechnology Information; https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 3 September 2019) and from the UNITE website (https://unite.ut.ee/ [27], accessed on 3 September 2019) was used to assess the similarity of obtained ITS sequences compared to the sequences in GenBank and UNITE public sequence databases. Sequences that remained unclassified at a kingdom or family level were discarded. The final criteria for a database match were as follows: query cover ≥80% and sequence similarity >92% (cut-off value at genus level) or sequence similarity ≥97–100% (cut-off at species level) [28,29].
2.4. Statistical Analyses
All statistical analyses were completed in R version 3.5.1 [32] using the community analysis specific package ‘vegan’ [33] and statistical packages ‘mvabund’ [34], ‘ggplot2’ [35], ‘multcomp’ [36], and ‘reshape2’ [37].
The correlation of phenological stages and sampling year with the composition of ectomycorrhizal community was tested with the ADONIS function (analogous for permutational multivariate analysis of variance (PERMANOVA)) in the vegan package [28] in R, which ran for 999 iterations using a Bray–Curtis distance matrix.
The differences in ectomycorrhizal community composition between phenological stages were visualized with a non-metric multidimensional scaling analysis (NMDS) ordination using the Bray–Curtis distance matrix.
Generalized linear models (GLMs) with Poisson regression were used to examine the effect of phenological stages on abundances of individual ectomycorrhizal taxa. For multiple comparisons between the time periods of each GLM, general linear hypotheses (GLHT) with Tukey contrast were used [33]. For taxa abundance comparison between time periods, when separated by sampling year, taxa that had an abundance higher than 2% were included for analysis.
A two-way ANOVA test was used to examine differences in the ratio of explorational types between phenological stages and sampled years. The homogeneity of variance assumptions was checked with a Levene test.
3. Results
3.1. Ectomycorrhizal Community Composition Changes along Transition between Phenological Stages
The environmental conditions in the previous month (PERMANOVA, p = 0.002) and year (PERMANOVA, p = 0.003) significantly affected ectomycorrhizal community composition in the month and year of sampling.
In sampling year 2016, the time between the phenological stages of dormancy and bud bursting was 9 weeks, and between bud bursting and full needle expansion this was 4 weeks. In sampling year 2017, the time between the phenological stages of dormancy and bud bursting was 5 weeks, and between bud bursting and full needle expansion this was 4 weeks.
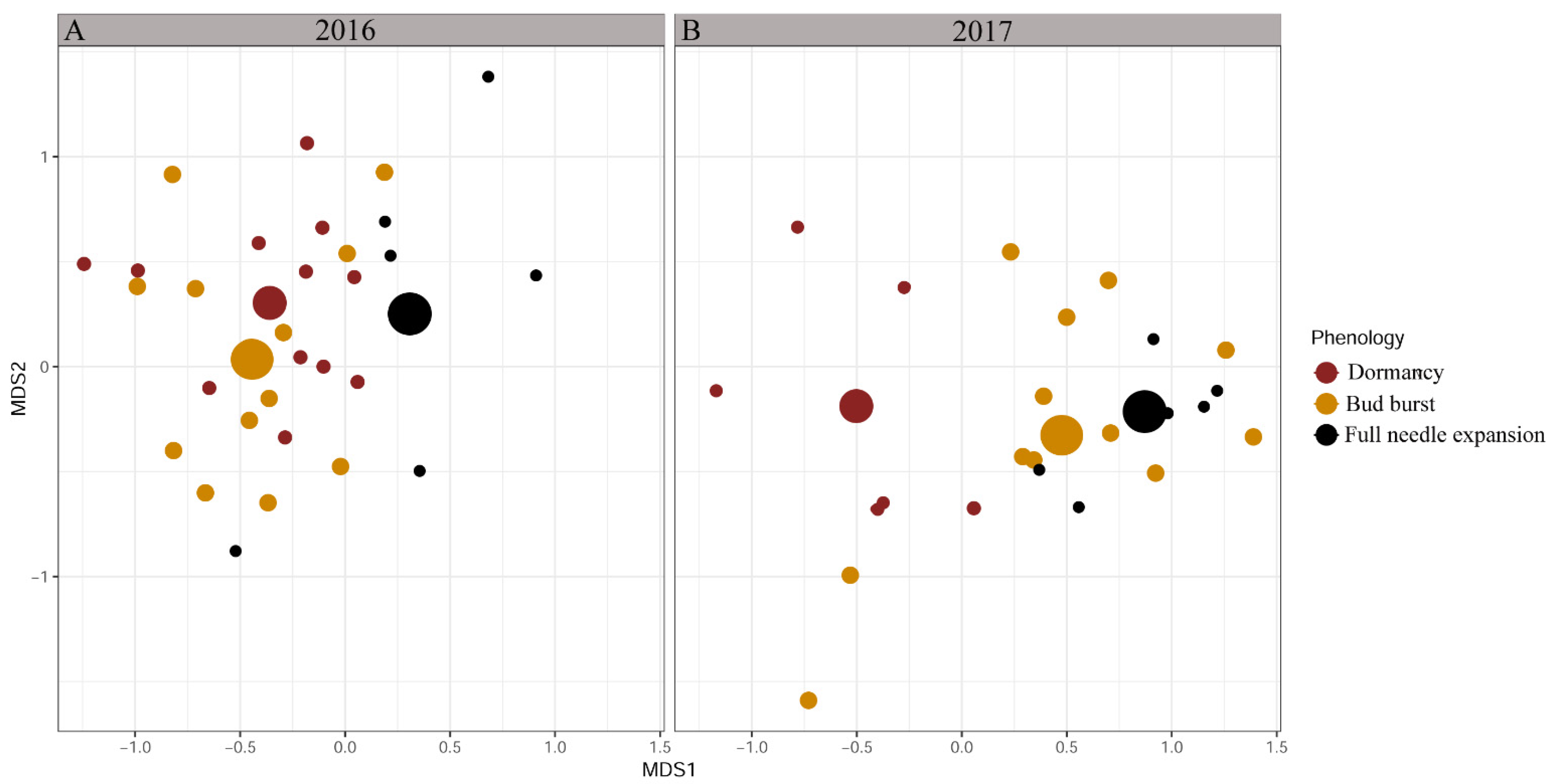
For sampling year 2016, NMDS showed a significantly different ectomycorrhizal community of bud bursting silver fir trees compared to dormant trees (PERMANOVA, p = 0.005) and trees with fully expanded needles (PERMANOVA, p = 0.006), meanwhile, there was an overlap in ectomycorrhizal community between dormant trees and trees with fully expanded needles (PERMANOVA, p = 0.080) (Figure 1A). A similar pattern was observed for sampling year 2017, where the ectomycorrhizal community of bud-bursting trees significantly differed from dormant trees (PERMANOVA, p = 0.012) but not from trees with fully expanded needles (PERMANOVA, p = 0.305). In contrast to sampling year 2016, in 2017 significant differences in ectomycorrhizal community between dormant trees and trees with fully expanded needles were observed (PERMANOVA, p = 0.022) (Figure 1B).
3.2. Individual Responses of Ectomycorrhizal OTUs along Transition between Phenological Stages
Ectomycorrhizal morphotyping of vital root tips combined with ITS sequencing retrieved 35 distinct ectomycorrhizal fungal OTUs belonging to 12 different genera, among which the most abundant were Tomentella, Russula, Lactarius, and Cenococcum; together, these represented 60.8% of all sequences. For a general view, root tips numbers of individually identified fungi per each phenological stage are represented in Table S2.
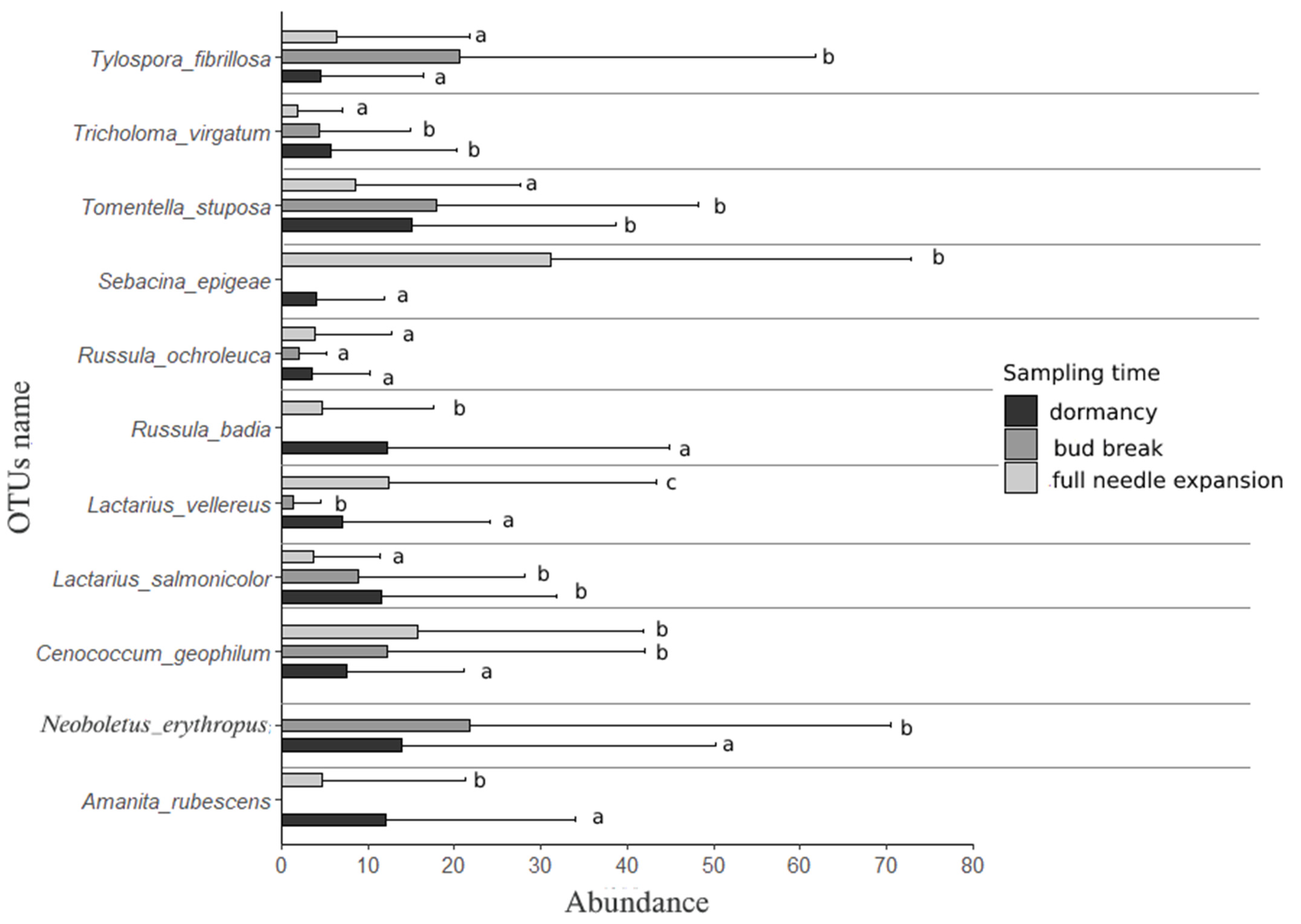
We observed 18 distinct ectomycorrhizal OTUs differing between three different time periods in the two years of sampling. Changes in ectomycorrhizal OTUs abundances along the transition between time periods were analyzed separately per sampling year. In sampling year 2016, dormant silver fir trees were associated with a significantly higher abundance of Amanita rubescens Pers. and Russula badia Beeli. In contrast, bud-bursting trees were significantly associated with Neoboletus erythropus (Pers.) C. Hahn and Tylospora fibrillosa (Burt) Donk, while we observed a higher abundance of Sebacina epigaea (Berk. and Broome) Neuhoff and Lactarius vellereus (Fr.) Fr. in association with a fully expanded needle phenological stage. Lactarius salmonicolor R. Heim. and Leclair, Tomentella stuposa (Link) Stalpers, and Tricholoma virgatum (Fr.) Kumm. were associated equally with dormant and bud-bursting trees and with a significantly lower abundance in the phenological stage of full needle expansion. Cenococcum geophilum Fr. had the lowest abundance during dormancy in 2016 while Russula ochroleuca Pers. had comparable abundance in all three phenological stages (Figure 2).
Significant differences in ectomycorrhizal OTUs abundances along the transition between time periods were further confirmed for sampling year 2017.
Dormant trees were associated significantly with Elaphomyces granulatus Fr., Lactarius lignyotus Fr., Russula badia Beeli, and Tricholoma virgatum (Fr.) Kumm. In contrast, bud-bursting trees were associated with Russula illota Romagn., Russula ochroleuca Pers., Russula sp. Pers., Tomentella sublilacina (Ellis and Holw.) Wakef., and Tylospora fibrillosa (Burt) Donk. Moreover, Lactarius tabidus Fr. and Amphinema byssoides (Pers.) J. Erikss. were the only ectomycorrhizal OTUs that were significantly associated with trees with fully expanded needles. (Figure 3).
4. Discussion
Using sequential soil coring and ectomycorrhizal community assessment, 35 different ectomycorrhizal OTUs were identified during three different time periods. The number of identified OTUs is comparable to other published studies in which authors used Sangar sequencing for identifying ectomycorrhizal symbionts of silver fir (varies from 15–42) [1,2,38,39].
By comparing ectomycorrhizal community composition between two sampling years and through three different time periods, which were linked to tree phenology, significant shifts in the community were confirmed. As some authors have already suggested regarding the mycorrhizal effect on host phenology [11,12,13], we tried to link observed shifts in ectomycorrhizal community composition with silver fir tree phenology, suggesting a possible functional role of ectomycorrhizal fungal species during different phenological stages, especially during the carbon expensive stage, such as bud break. However, other factors, such as fine root dynamic and environmental conditions, were also taken into consideration.
In 2016, the ectomycorrhizal community of bud-bursting trees significantly differed from dormant trees and from trees with needles that were fully expanded. However, there was an overlap between the ectomycorrhizal community of dormant trees and trees with full needle expansion. Similar patterns were observed in 2017, where the ectomycorrhizal community of dormant trees significantly differed from bud-bursting trees as well as from trees with needles that were fully expanded. In sampling year 2017, we observed a dominance of different ectomycorrhizal fungi compared to 2016, which could be due to the dry winter in 2016 and the overall drier year, with a smaller amount of precipitation compared to sampling year 2017 [40].
Overall, through observed findings we confirmed the expected shift in the ectomycorrhizal community between phenological stages. However, in contrast to the findings of Hupperts et al. [41], we did not confirm statistically significant differences in the ectomycorrhizal community between all three phenological stages. Namely, there was an overlap either between dormant trees and trees with fully expanded needles (2016) or between bud-bursting trees and trees with fully expanded needles (2017).
Estimating the role of ectomycorrhizal root tips in nutrient and carbon cycling requires a quantification of their numbers and turnover rates [42]. Observed differences in ectomycorrhizal OTUs abundance between the phenological stages in our study could thereby be a result of the production of new root tips and/or their turnover. The mortality of fine roots is greater during winter [43] as root growth declines substantially over winter when water requirements are at their lowest [44]. In contrast, the initiation of fine root growth in the forest floor coincides with leaf expansion in the forest canopy, and is most rapid in early summer [45]. A significant difference in ectomycorrhizal community composition during bud break and full needle expansion in sampling year 2017 could thereby be linked with fine root seasonal dynamics. However, an overlap between dormancy and full needle expansion in sampling year 2016 is difficult to explain through only fine root dynamics.
Although there have been several attempts to link ectomycorrhizal root tip turnover rates to needle or leaf biomass [42], so far, there are no published data about the possible relation between ectomycorrhizal root tip turnover and needle/leaf phenology. However, due to a relatively long ectomycorrhizal root tips’ lifespan (minimum reported is 129 days and maximum is 675 days) [46,47,48,49,50], the turnover rate in our study can hardly be linked to only fine root dynamics and ectomycorrhizal root tip turnover. Thus, the time-lapse between two sampling periods may depend partly on fine root seasonal dynamics and ectomycorrhizal root tip turnover; other factors, such as host tree phenology, should also be taken into consideration.
The observed changes in the ectomycorrhizal community between dormant and bud-bursting silver fir trees can thereby be linked to the functional role of ectomycorrhizal fungal species. As already reported, ectomycorrhizal symbiosis can help host plants through a timely provision of carbon supply, particularly when demand is high [11,51,52]. Several studies have pointed to the possible degradative abilities of ectomycorrhizal fungi [53,54], which may have retained and can, under specific conditions, express genes for decomposing enzymes [53,55,56,57]. For members of genera Tomentella and Tylospora, Read and Perez-Moreno [58] suggested the species’ ability to decompose carbohydrates. For ectomycorrhizal fungi Tylospora fibrillosa, the presence of manganese peroxidase, which is one of the key enzymes responsible for lignin degradation, was confirmed in the study by Cairney et al. [59] as well as in the study by Burke and Cairney [59]. Moreover, Chambers et al. [60] confirmed the extracellular activity of manganese peroxidase in part of a gene identified in the ectomycorrhizal fungus Tylospora fibrillosa, which additionally supports the records about capable production of manganese peroxidase by this ectomycorrhizal fungus. Along with Tylospora fibrillosa, for members of genera Russula, the presence of laccase enzymes has already been confirmed [56,57,59]. In our study, ectomycorrhizal fungi Russula illota, Russula sp., Russula ochroleuca, Tomentella sublilacina, and Tylospora fibrillosa were the most abundant ectomycorrhizal fungi associated with bud-bursting trees, with significantly higher abundances during bud burst. Thereby, we could potentially link higher abundances of the mentioned species to their functional role and possible degradative abilities during carbon-expensive phenophase, such as bud-brake. In contrast, Hupperts et al. [41] confirmed only higher activity of β-glucosidase during dormancy and a weak positive relation between host phenology and overall ectomycorrhizal community enzymatic activity. Although several ectomycorrhizal fungi are predicted to be able to decompose organic matter, we must take into consideration that they are not able to survive solely as saprotrophs as their ability to decompose lignocellulose and degrade cellulose is much lower compared to wood-decaying fungi and/or litter saprotrophs and insufficient for growth [61].
Among others, the observed ectomycorrhizal fungal occurrence pattern and differences in overall ectomycorrhizal communities between phenological stages could be linked with seasonal variations and the change in environmental conditions. There are quite a few studies in which authors confirmed the significant impact of seasonal variations on ectomycorrhizal community compositional changes [11,14,62,63,64,65]; however, direct comparison of our results to published ones would not be relevant due to the upgraded soil sampling in our study, which was performed more frequently (at a monthly basis), rather than once per individual season. Precipitation, soil moisture, and temperature are some of the most important environmental variables affecting ectomycorrhizal community composition and richness [66,67,68,69]. The significant impact of precipitation on the fungal community was also confirmed by Hawkes et al. [70], who observed a shift in fungal community in the months when the rainfall during the month before sampling was extremely low. Based on our previously published study within a PhD thesis [40], observed community shifts could thereby be further linked with monthly precipitation. In sampling year 2016, a lower amount of precipitation was recorded in April compared to February and May (Table S3); therefore, a significantly different ectomycorrhizal community during bud break (May) compared to dormancy (March) and full needle expansion (June) could be a result of a change in precipitation. However, in sampling year 2017, a lower amount of precipitation was recorded for February and March; therefore, shifts in the community during dormancy (March) cannot be exclusively linked with precipitation as a similar community was observed during bud break (April) and full needle expansion (May). Shifts in the ectomycorrhizal community could also be reflected by significant changes in temperature [71,72]; however, in our PhD thesis [40], significant changes in temperature were not confirmed.
A possible mechanism for temporal shifts in the ectomycorrhizal community could thereby be a combination of various biotic and abiotic factors, e.g., host trees’ physiology, root turnover, ectomycorrhizal lifespan, and environmental conditions.
5. Conclusions
In this study, we found statistically significant ectomycorrhizal community compositional shifts between three different time periods linked to silver fir needle phenology. Moreover, we recorded significant differences in ectomycorrhizal fungal OTUs abundances associated with different time periods by which we indicated a potential link between ectomycorrhizal community composition and host tree phenology, especially during bud-burst, which is known to be a highly C-demanding growth phase. The observed results most likely reflect both the influence of host tree physiology and the change in environmental conditions. However, further analyses of ectomycorrhizal fungal enzymatic activities are needed to link shifts in ectomycorrhizal community composition with host trees’ phenology and for a better understanding of the functional ecology of ectomycorrhizal fungal symbionts.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/f12121719/s1. Table S1: General linear hypothesis (GLHT) with Tukey contrast based on generalized linear models (GLMs) with Poisson regression of selected ectomycorrhizal taxa (n.s = p > 0.05; p < 0.001 ***; p < 0.01 **; p < 0.05 *). Ectomycorrhizal taxa with abundances lower than 2% were excluded from statistical analyses; ‘absent’ in the table means that the specified ECM taxa did not appear in one of two compared phenological stages, therefore, comparison with GLHT was not possible (for detailed information look at Table S2). Table S2: Ectomycorrhizal taxa presence (+) or absence (−) during individual phenological stage with number of root tips, separated by sampling year (2016, 2017). Table S3: Monthly precipitation per sampling time and sampling year.
Author Contributions
T.U.N., H.K. and T.G. conceived and designed the experiments; T.U.N. and R.D. performed the experiments; T.U.N. and R.D. analyzed the data; T.U.N. wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
The study was funded by the Young Researcher Scheme (Slovenian Research Agency) and co-financed by The Research Program P4-0107 Forest Biology, Ecology, and Technology (Slovenian Research Agency) and the LIFEGENMON project (LIFE ENV/SI/000148).
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available in the GenBank repository, under accession numbers MK820070-MK820175.
Acknowledgments
The authors acknowledge help during fieldwork from Miran Praznik (Slovenia Forest Service, Regional Unit Maribor) and lab assistance from Barbara Štupar and Melita Hrenko (Slovenian Forestry Institute). Anonymous reviewers are acknowledged for their contribution to improvement in the manuscript in the revision process.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ważny, R. Ectomycorrhizal communities associated with silver fir seedlings (Abies alba Mill.) differ largely in mature silver fir stands and in Scots pine forecrops. Ann. For. Sci. 2014, 71, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Rudawska, M.; Pietras, M.; Smutek, I.; Strzeliński, P.; Leski, T. Ectomycorrhizal fungal assemblages of Abies alba Mill. outside its native range in Poland. Mycorrhiza 2016, 26, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Vitasse, Y.; Delzon, S.; Bresson, C.C.; Michalet, R.; Kremer, A. Altitudinal differentiation in growth and phenology among populations of temperate-zone tree species growing in a common garden. Can. J. For. Res. 2009, 39, 1259–1269. [Google Scholar] [CrossRef] [Green Version]
- Vitasse, Y.; Delzon, S.; Dufrêne, E.; Pontailler, J.Y.; Louvet, J.M.; Kremer, A.; Michalet, R. Leaf phenology sensitivity to temperature in European trees: Do within-species populations exhibit similar responses? Agric. For. Meteorol. 2009, 149, 735–744. [Google Scholar] [CrossRef]
- Arend, M.; Brem, A.; Kuster, T.M.; Günthardt-Goerg, M.S. Seasonal photosynthetic responses of European oaks to drought and elevated daytime temperature. Plant Biol. 2013, 15, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Delpierre, N.; Vitasse, Y.; Chuine, I.; Guillemot, J.; Bazot, S.; Rutishauser, T.; Rathgeber, C.B.K. Temperate and boreal forest tree phenology: From organ-scale processes to terrestrial ecosystem models. Ann. For. Sci. 2016, 73, 5–25. [Google Scholar] [CrossRef] [Green Version]
- Dickie, I.A.; Montgomery, R.A.; Reich, P.B.; Schnitzer, S.A. Physiological and phenological responses of oak seedlings to oak forest soil in the absence of trees. Tree Physiol. 2007, 27, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Unuk, T.; Martinović, T.; Finžgar, D.; Šibanc, N.; Grebenc, T.; Kraigher, H. Root-associated fungal communities from two phenologically contrasting silver fir (Abies alba mill.) groups of trees. Front. Plant Sci. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, B.Y.E.B.; Allen, M.E. Water relations of xeric grasses in the field: Interactions of mycorrhizas and competition. New Phytol. 1986, 104, 559–571. [Google Scholar] [CrossRef]
- Garbaye, J. Garbaye_citation. In Mycorrhizae: Physiology and Genetics-Les Mycorrhizes: Physiologie et Genetique; Gianinazzi-Pearson, V., Gianinazzi, S., Eds.; INRA: Paris, France, 1986; pp. 493–496. [Google Scholar]
- Courty, P.E.; Bréda, N.; Garbaye, J. Relation between oak tree phenology and the secretion of organic matter degrading enzymes by Lactarius quietus ectomycorrhizas before and during bud break. Soil Biol. Biochem. 2007, 39, 1655–1663. [Google Scholar] [CrossRef]
- Waldrop, M.P.; Firestone, M.K. Seasonal dynamics of microbial community composition and function in oak canopy and open grassland soils. Microb. Ecol. 2006, 52, 470–479. [Google Scholar] [CrossRef]
- Dhuli, P.; Rohloff, J.; Strimbeck, G.R. Metabolite changes in conifer buds and needles during forced bud break in Norway spruce (Picea abies) and European silver fir (Abies alba). Front. Plant Sci. 2014, 5, 706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, J.F.; Miller, O.K.; Horton, J.L. Seasonal dynamics of ectomycorrhizal fungus assemblages on oak seedlings in the southeastern Appalachian Mountains. Mycorrhiza 2008, 18, 123–132. [Google Scholar] [CrossRef]
- Bajc, M.; Aravanopoulos, F.; Westergren, M.; Fussi, B.; Kavaliauskas, D.; Alizoti, P.; Kraigher, H. (Eds.) Manual for Forest Genetic Monitoring; Silva Slovenica Publishing Centre: Ljubljana, Slovenia, 2020. [Google Scholar]
- Kraigher, H. Tipi ektomikorize-taksonomija, pomen in aplikacihe-Types of ectomycorrhizae-their taxonomy, role and applocation. Acta Silvae Ligni 1996, 49, 33–66. [Google Scholar]
- Taylor, D.L.; Bruns, T.D. Community structure of ectomycorrhizal fungi in a Pinus muricata forest: Minimal overlap between the mature forest and resistant propagule communities. Mol. Ecol. 1999, 8, 1837–1850. [Google Scholar] [CrossRef] [Green Version]
- Baar, J.; Horton, T.R.; Kretzer, A.M.; Bruns, T.D. Mycorrhizal colonization of Pinus muricata from resistant propagules after a stand-replacing wildfire. New Phytol. 1999, 143, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Železnik, P.; Hrenko, M.; Then, C.; Koch, N.; Grebenc, T.; Levanič, T.; Kraigher, H. CASIROZ: Root parameters and types of ectomycorrhiza of young beech plants exposed to different ozone and light regimes. Plant Biol. 2007, 9, 298–308. [Google Scholar] [CrossRef]
- Chen, W.; Koide, R.T.; Adams, T.S.; DeForest, J.L.; Cheng, L.; Eissenstat, D.M. Root morphology and mycorrhizal symbioses together shape nutrient foraging strategies of temperate trees. Proc. Natl. Acad. Sci. USA 2016, 113, 8741–8746. [Google Scholar] [CrossRef] [Green Version]
- Železnik, P.; Vilhar, U.; Starr, M.; de Groot, M.; Kraigher, H. Fine root dynamics in Slovenian beech forests in relation to soil temperature and water availability. Trees Struct. Funct. 2016, 30, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Agerer, R. Characterization of Ectomycorrhiza. Methods Microbiol. 1991, 23, 25–73. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal rna genes for phylogenetics. In PCR Protocols; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. [Google Scholar]
- Sulzbacher, M.A.; Grebenc, T.; Cabral, T.S.; Giachini, A.J.; Goto, B.T.; Smith, M.E.; Baseia, I.G. Restingomyces, a new sequestrate genus from the Brazilian Atlantic rainforest that is phylogenetically related to early-diverging taxa in Trappeaceae (Phallales). Mycologia 2016, 108, 954–966. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef] [PubMed]
- Porras-Alfaro, A.; Liu, K.L.; Kuske, C.R.; Xiec, G. From genus to phylum: Large-subunit and internal transcribed spacer rRNA operon regions show similar classification accuracies influenced by database composition. Appl. Environ. Microbiol. 2014, 80, 829–840. [Google Scholar] [CrossRef] [Green Version]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal Identification Using Molecular Tools: A Primer for the Natural Products Research Community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef]
- Rinaldi, A.C.; Comandini, O.; Kuyper, T.W. Ectomycorrhizal fungal diversity: Separating the wheat from the chaff. Fungal Divers. 2008, 33, 1–45. [Google Scholar]
- Tedersoo, L.; May, T.W.; Smith, M.E. Ectomycorrhizal lifestyle in fungi: Global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 2010, 20, 217–263. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2016. [Google Scholar]
- Oksanen, J.F.; Blanchet, G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2018. Available online: https://github.com/vegandevs/vegan (accessed on 3 September 2019).
- Wang, Y.; Naumann, U.; Wright, S.T.; Warton, D.I. Mvabund—An R package for model-based analysis of multivariate abundance data. Methods Ecol. Evol. 2012, 3, 471–474. [Google Scholar] [CrossRef]
- Wickham, H. Package ‘ggplot2’: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Reshaping data with the reshape package. J. Stat. Softw. 2007, 21, 1–20. [Google Scholar] [CrossRef]
- Cremer, E. Population genetics of silver fir (Abies alba Mill.) in the Northern Black Forest–preconditions for the recolonization of windthrow areas and associated ectomycorrhizal communities. Phillips-Universität Marbg. 2009, 1, 103. [Google Scholar] [CrossRef]
- Schirkonyer, U.; Bauer, C.; Rothe, G.M. Ectomycorrhizal diversity at five different tree species in forests of the Taunus Mountains in Central Germany. Open J. Ecol. 2013, 3, 66–81. [Google Scholar] [CrossRef] [Green Version]
- Unuk Nahberger, T. Ektomikorizni Simbionti Bele Jelke (Abies alba Mill.); Univerza v Ljubljani, Biotehniška Fakulteta: Ljubljana, Slovenia, 2020. [Google Scholar]
- Hupperts, S.F.; Karst, J.; Pritsch, K.; Landhäusser, S.M. Host phenology and potential saprotrophism of ectomycorrhizal fungi in the boreal forest. Funct. Ecol. 2017, 31, 116–126. [Google Scholar] [CrossRef]
- Helmisaari, H.-S.; Ostonen, I.; Lohmus, K.; Derome, J.; Lindroos, A.-J.; Merila, P.; Nojd, P. Ectomycorrhizal root tips in relation to site and stand characteristics in Norway spruce and Scots pine stands in boreal forests. Tree Physiol. 2009, 29, 445–456. [Google Scholar] [CrossRef] [Green Version]
- Tierney, G.L.; Fahey, T.J.; Groffman, P.M.; Hardy, J.P.; Fitzhugh, R.D.; Driscoll, C.T. Soil freezing alters fine root dynamics in a northern hardwood forest. Biogeochemistry 2001, 56, 175–190. [Google Scholar] [CrossRef]
- Hendrick, R.L.; Pregitzer, K.S. Temporal and Depth-Related Patterns of Fine Root Dynamics in Northern Hardwood Forests. J. Ecol. 1996, 84, 167. [Google Scholar] [CrossRef]
- Fahey, T.J.; Hughes, J.W. Fine Root Dynamics in a Northern Hardwood Forest Ecosystem, Hubbard Brook Experimental Forest, NH. J. Ecol. 1994, 82, 533. [Google Scholar] [CrossRef]
- Harley, J.L. Ecology of ectotrophic mycorrhizas. In The Biology of Mycorrhiza; Polunin, N., Ed.; Leonard Hill: London, UK, 1969; pp. 150–162. [Google Scholar]
- Rygiewicz, P.T.; Johnson, M.G.; Ganio, L.M.; Tingey, D.T.; Storm, M.J. Lifetime and temporal occurrence of ectomycorrhizae on ponderosa pine (Pinus ponderosa Laws.) seedlings grown under varied atmospheric CO2 and nitrogen levels. Plant Soil 1997, 189, 275–287. [Google Scholar] [CrossRef]
- Majdi, H.; Damm, E.; Nylund, J.-E. Longevity of mycorrhizal roots depends on branching order and nutrient availability. New Phytol. 2001, 150, 195–202. [Google Scholar] [CrossRef]
- Fernandez, C.W.; McCormack, M.L.; Hill, J.M.; Pritchard, S.G.; Koide, R.T. On the persistence of Cenococcum geophilum ectomycorrhizas and its implications for forest carbon and nutrient cycles. Soil Biol. Biochem. 2013, 65, 141–143. [Google Scholar] [CrossRef]
- Kou, L.; McCormack, M.L.; Chen, W.; Guo, D.; Wang, H.; Gao, W.; Yang, H.; Li, S. Nitrogen ion form and spatio-temporal variation in root distribution mediate nitrogen effects on lifespan of ectomycorrhizal roots. Plant Soil 2017, 411, 261–273. [Google Scholar] [CrossRef]
- Cullings, K.; Ishkhanova, G.; Henson, J. Defoliation effects on enzyme activities of the ectomycorrhizal fungus Suillus granulatus in a Pinus contorta (lodgepole pine) stand in Yellowstone National Park. Oecologia 2008, 158, 77–83. [Google Scholar] [CrossRef]
- Talbot, J.M.; Allison, S.D.; Treseder, K.K. Decomposers in disguise: Mycorrhizal fungi as regulators of soil C dynamics in ecosystems under global change. Funct. Ecol. 2008, 22, 955–963. [Google Scholar] [CrossRef] [Green Version]
- Bödeker, I.T.M.; Nygren, C.M.R.; Taylor, A.F.S.; Olson, Å.; Lindahl, B.D. ClassII peroxidase-encoding genes are present in a phylogenetically wide range of ectomycorrhizal fungi. ISME J. 2009, 3, 1387–1395. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, B.D.; Tunlid, A. Ectomycorrhizal fungi-potential organic matter decomposers, yet not saprotrophs. New Phytol. 2015, 205, 1443–1447. [Google Scholar] [CrossRef] [PubMed]
- Hibbett, D.S.; Gilbert, L.; Donoghue, M.J. Evolutionary instability of ectomycorrhizal symbioses in basidiomycetes. Nat. Cell Biol. 2000, 407, 506–508. [Google Scholar] [CrossRef]
- Chen, D.M.; Taylor, A.F.S.; Burke, R.M.; Cairney, J.W.G. Identification of genes for lignin peroxidases and manganese peroxidases in ectomycorrhizal fungi. New Phytol. 2001, 152, 151–158. [Google Scholar] [CrossRef]
- Chen, D.M.; Bastias, B.A.; Taylor, A.F.S.; Cairney, J.W.G. Identification of laccase-like genes in ectomycorrhizal basidiomycetes and transcriptional regulation by nitrogen in Piloderma byssinum. New Phytol. 2003. [Google Scholar] [CrossRef]
- Read, D.J.; Perez-Moreno, J. Mycorrhizas and nutrient cycling in ecosystems-A journey towards relevance? New Phytol. 2003, 157, 475–492. [Google Scholar] [CrossRef]
- Burke, R.M.; Cairney, J.W.G. Laccases and other polyphenol oxidases in ecto- and ericoid mycorrhizal fungi. Mycorrhiza 2002, 12, 105–116. [Google Scholar] [CrossRef]
- Chambers, S.M.; Burke, R.M.; Brooks, P.R.; Cairney, J.W.G. Molecular and biochemical evidence for manganese-dependent peroxidase activity in Tylospora fibrillosa. Mycol. Res. 1999, 103, 1098–1102. [Google Scholar] [CrossRef]
- Martin, F.; Kohler, A.; Murat, C.; Veneault-Fourrey, C.; Hibbett, D.S. Unearthing the roots of ectomycorrhizal symbioses. Nat. Rev. Microbiol. 2016, 14, 760–773. [Google Scholar] [CrossRef]
- Koide, R.T.; Shumway, D.L.; Xu, B.; Sharda, J.N. On temporal partitioning of a community of ectomycorrhizal fungi. New Phytol. 2007, 174, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Aponte, C.; Marañón, T.; García, L.V. Microbial C, N and P in soils of Mediterranean oak forests: Influence of season, canopy cover and soil depth. Biogeochemistry 2010, 101, 77–92. [Google Scholar] [CrossRef] [Green Version]
- Richard, F.; Roy, M.; Shahin, O.; Sthultz, C.; Duchemin, M.; Joffre, R.; Selosse, M.A. Ectomycorrhizal communities in a Mediterranean forest ecosystem dominated by Quercus ilex: Seasonal dynamics and response to drought in the surface organic horizon. Ann. For. Sci. 2011, 68, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Vořiškova, J.; Brabcová, V.; Cajthaml, T.; Baldrian, P. Seasonal dynamics of fungal communities in a temperate oak forest soil. New Phytol. 2014, 201, 269–278. [Google Scholar] [CrossRef]
- Chen, M.M.; Zhu, Y.G.; Su, Y.H.; Chen, B.D.; Fu, B.J.; Marschner, P. Effects of soil moisture and plant interactions on the soil microbial community structure. Eur. J. Soil Biol. 2007, 43, 31–38. [Google Scholar] [CrossRef]
- Schimel, J.P.; Gulledge, J.M.; Clein-Curley, J.S.; Lindstrom, J.E.; Braddock, J.F. Moisture effects on microbial activity and community structure in decomposing birch litter in the Alaskan taiga. Soil Biol. Biochem. 1999, 31, S0038–S0717. [Google Scholar] [CrossRef]
- Williams, M.A. Response of microbial communities to water stress in irrigated and drought-prone tallgrass prairie soils. Soil Biol. Biochem. 2007, 39, 2750–2757. [Google Scholar] [CrossRef]
- Erlandson, S.R.; Savage, J.A.; Cavender-Bares, J.M.; Peay, K.G. Soil moisture and chemistry influence diversity of ectomycorrhizal fungal communities associating with willow along an hydrologic gradient. FEMS Microbiol. Ecol. 2016, 92, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkes, C.V.; Kivlin, S.N.; Rocca, J.D.; Huguet, V.; Thomsen, M.A.; Suttle, K.B. Fungal community responses to precipitation. Glob. Chang. Biol. 2011, 17, 1637–1645. [Google Scholar] [CrossRef]
- Castro, H.F.; Classen, A.T.; Austin, E.E.; Norby, R.J.; Schadt, C.W. Soil microbial community responses to multiple experimental climate change drivers. Appl. Environ. Microbiol. 2010, 76, 999–1007. [Google Scholar] [CrossRef] [Green Version]
- Bahram, M.; Põlme, S.; Kõljalg, U.; Zarre, S.; Tedersoo, L. Regional and local patterns of ectomycorrhizal fungal diversity and community structure along an altitudinal gradient in the Hyrcanian forests of northern Iran. New Phytol. 2012, 193, 465–473. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Non-metric, multidimensional scaling ordination (MDS) of ectomycorrhizal community based on phenological stages, calculated by Bray–Curtis distance, separated by sampled year. Presented are centroids of each group of samples. (A) In sampling year 2016, bud-bursting trees differ significantly from dormant trees (PERMANOVA, p = 0.005) and from trees with fully expanded needles (PERMANOVA, p = 0.006). (B) In sampling year 2017, significant differences in dormant trees compared to bud-bursting trees and trees with fully expanded needles were recorded (PERMANOVA, p = 0.012; p = 0.022).
Figure 1.
Non-metric, multidimensional scaling ordination (MDS) of ectomycorrhizal community based on phenological stages, calculated by Bray–Curtis distance, separated by sampled year. Presented are centroids of each group of samples. (A) In sampling year 2016, bud-bursting trees differ significantly from dormant trees (PERMANOVA, p = 0.005) and from trees with fully expanded needles (PERMANOVA, p = 0.006). (B) In sampling year 2017, significant differences in dormant trees compared to bud-bursting trees and trees with fully expanded needles were recorded (PERMANOVA, p = 0.012; p = 0.022).

Figure 2.
Mean and maximum abundances of selected ectomycorrhizal OTUs at species level in sampling year 2016. Ectomycorrhizal OTUs with abundances below 2% were excluded from the plot. Letters mark significantly different result.
Figure 2.
Mean and maximum abundances of selected ectomycorrhizal OTUs at species level in sampling year 2016. Ectomycorrhizal OTUs with abundances below 2% were excluded from the plot. Letters mark significantly different result.

Figure 3.
Mean and maximum abundance of selected ectomycorrhizal OTUs at species level in sampling year 2017. Ectomycorrhizal OTUs with abundances below 2% were excluded from the plot. Letters mark significantly different results.
Figure 3.
Mean and maximum abundance of selected ectomycorrhizal OTUs at species level in sampling year 2017. Ectomycorrhizal OTUs with abundances below 2% were excluded from the plot. Letters mark significantly different results.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Ectomycorrhizal sampling time according to individual phenological stage in sampling years 2016 and 2017.
Table 1.
Ectomycorrhizal sampling time according to individual phenological stage in sampling years 2016 and 2017.
| Phenophase|Sampling Time | Year 2016 | Year 2017 |
|---|---|---|
| Dormancy | 16 March | 20 March |
| Bud bursting | 16 May | 28 April |
| Full needle expansion | 15 June | 25 May |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Unuk Nahberger, T.; Damjanič, R.; Kraigher, H.; Grebenc, T. Potential Link between Ectomycorrhizal Community Composition and Host Tree Phenology. Forests 2021, 12, 1719. https://doi.org/10.3390/f12121719
AMA Style
Unuk Nahberger T, Damjanič R, Kraigher H, Grebenc T. Potential Link between Ectomycorrhizal Community Composition and Host Tree Phenology. Forests. 2021; 12(12):1719. https://doi.org/10.3390/f12121719
Chicago/Turabian StyleUnuk Nahberger, Tina, Rok Damjanič, Hojka Kraigher, and Tine Grebenc. 2021. "Potential Link between Ectomycorrhizal Community Composition and Host Tree Phenology" Forests 12, no. 12: 1719. https://doi.org/10.3390/f12121719
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.