
Surprising Drought Tolerance of Fir (Abies) Species between Past Climatic Adaptation and Future Projections Reveals New Chances for Adaptive Forest Management

 , , and
, , and 
Abstract
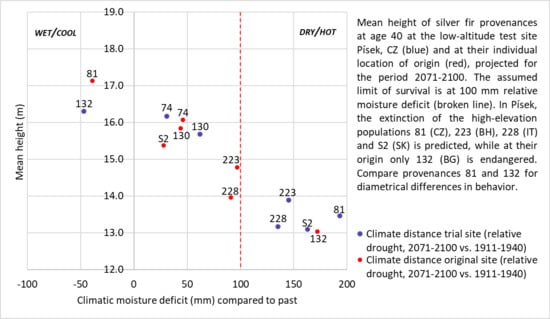
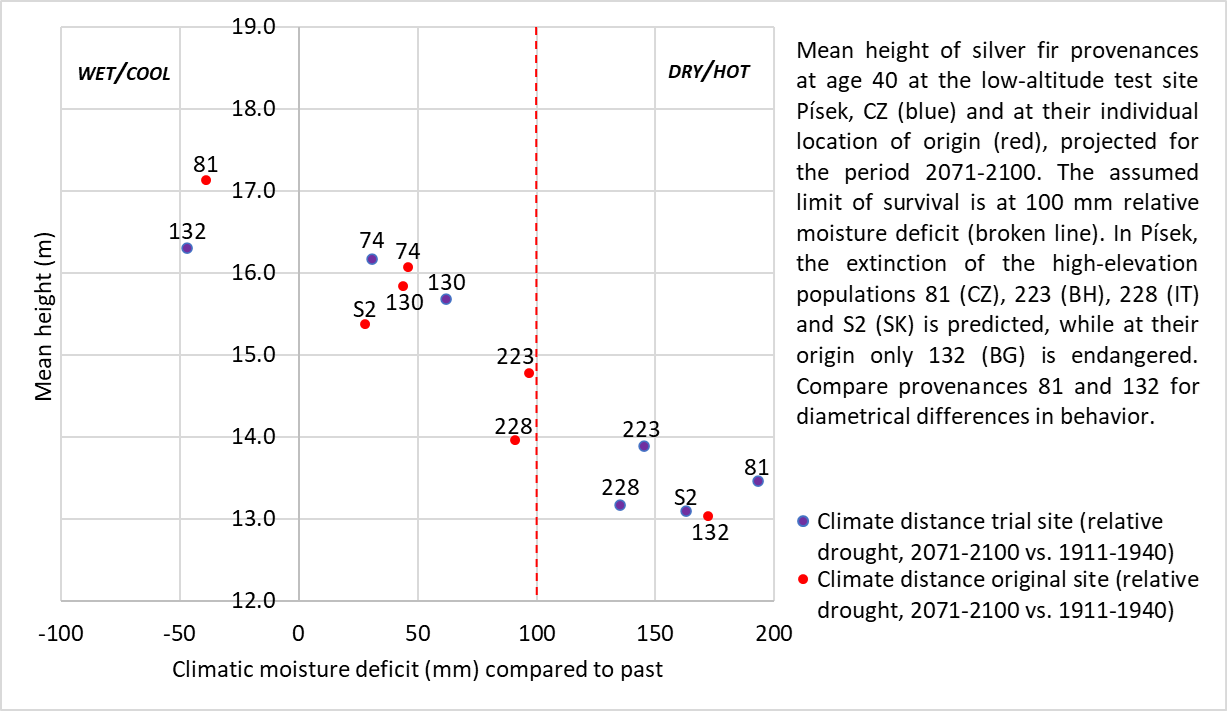
:
1. Introduction
2. Materials and Methods
2.1. Description of the Three Provenance Trials
2.2. Source of Data and Method of Analysis
3. Results
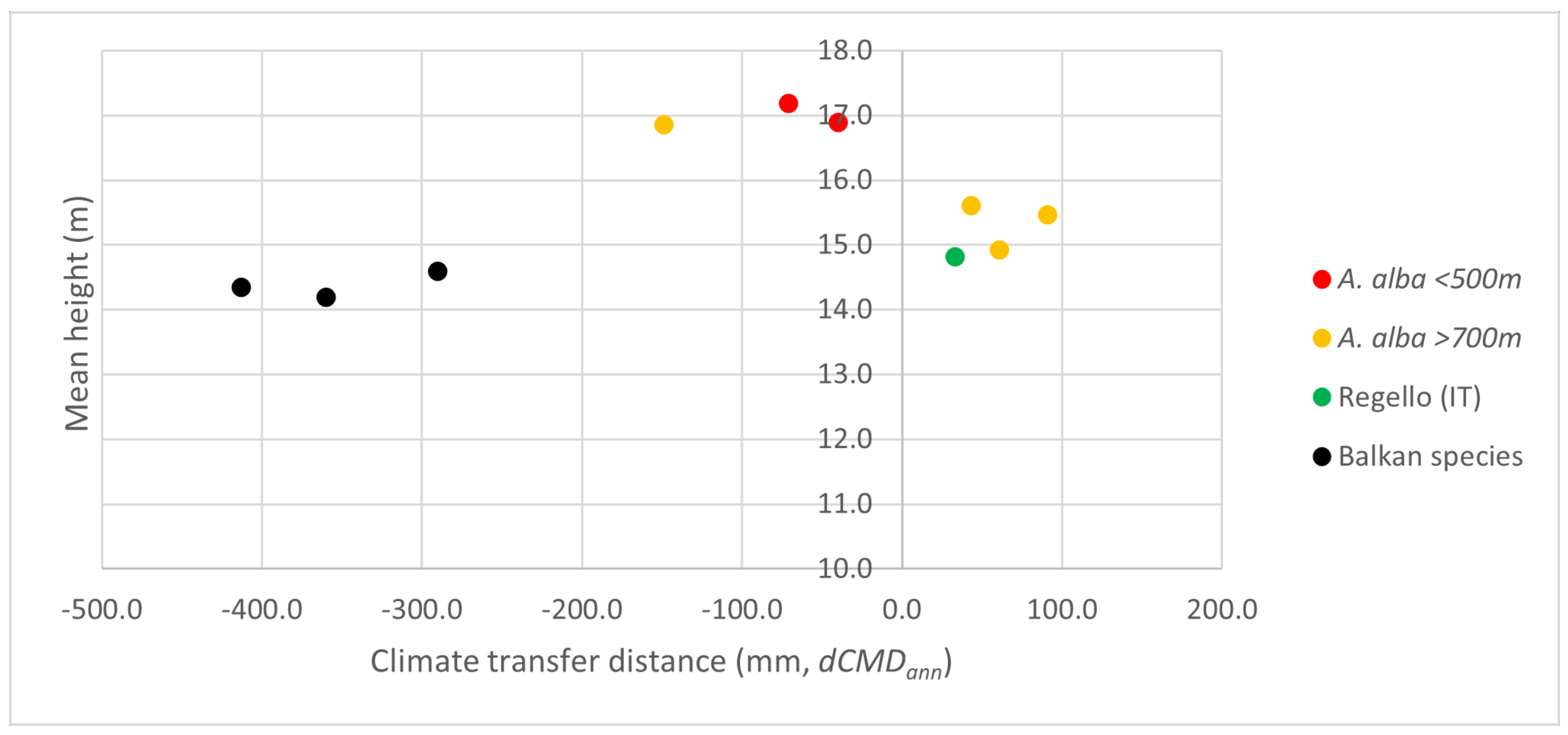
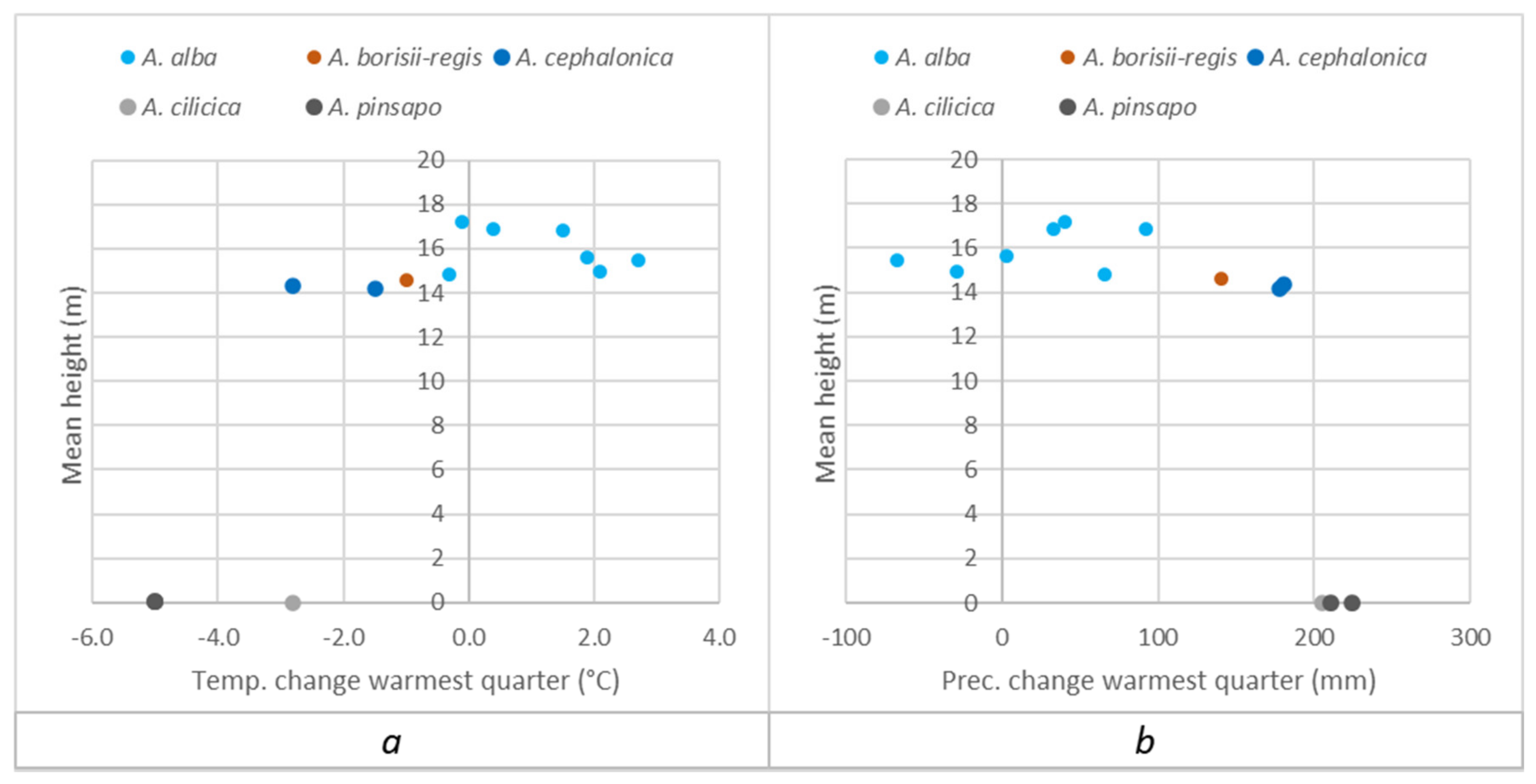
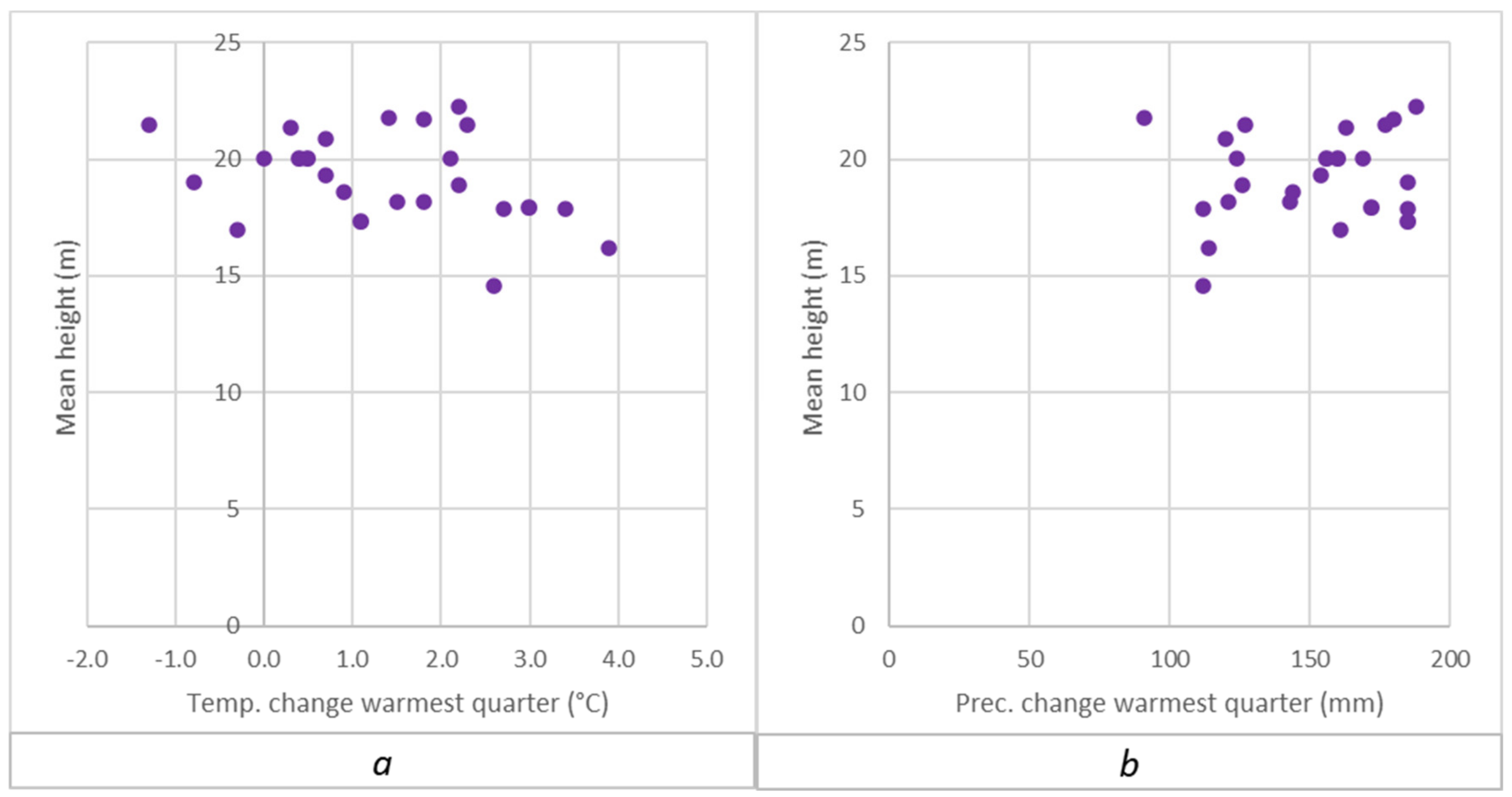
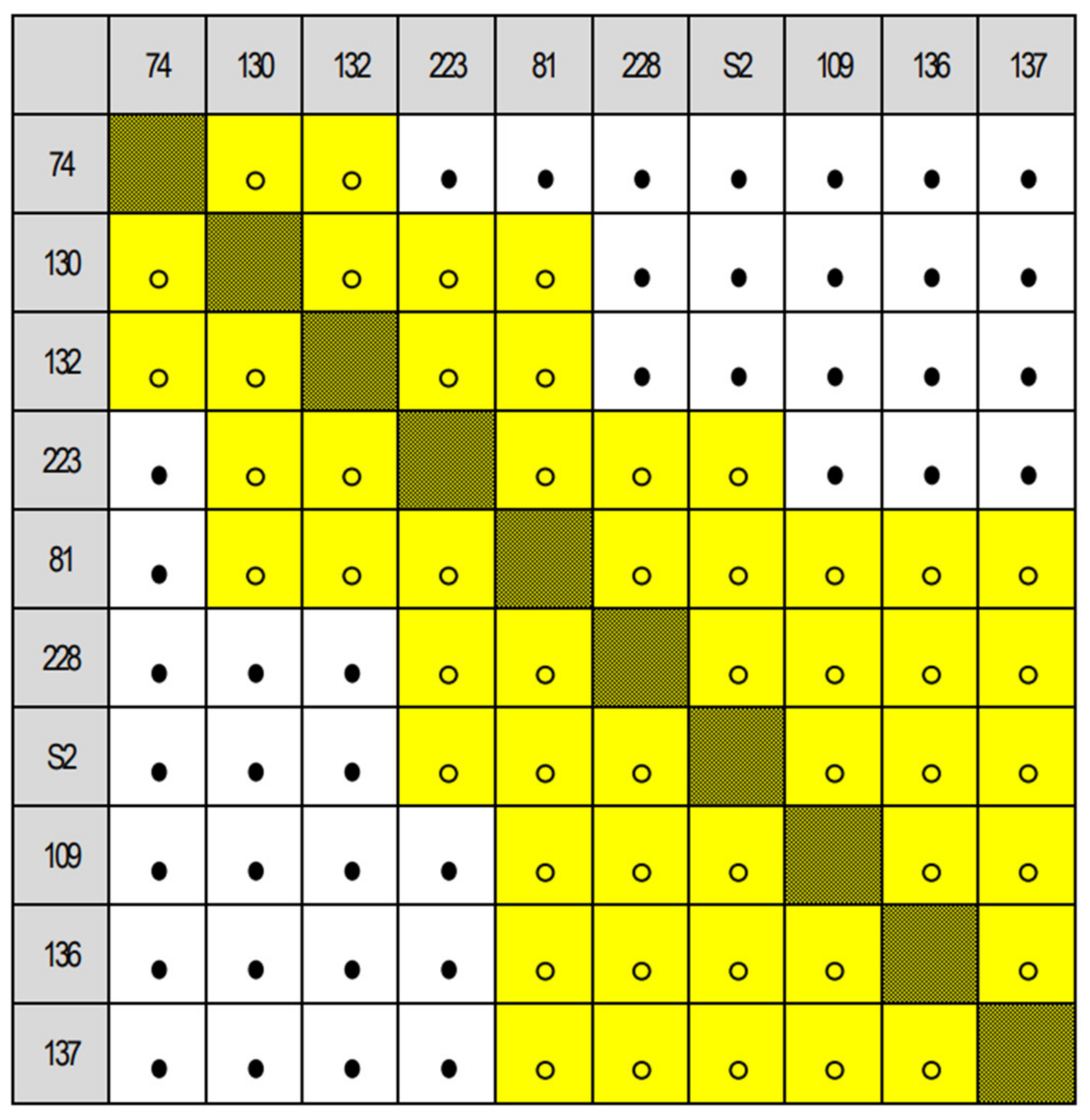
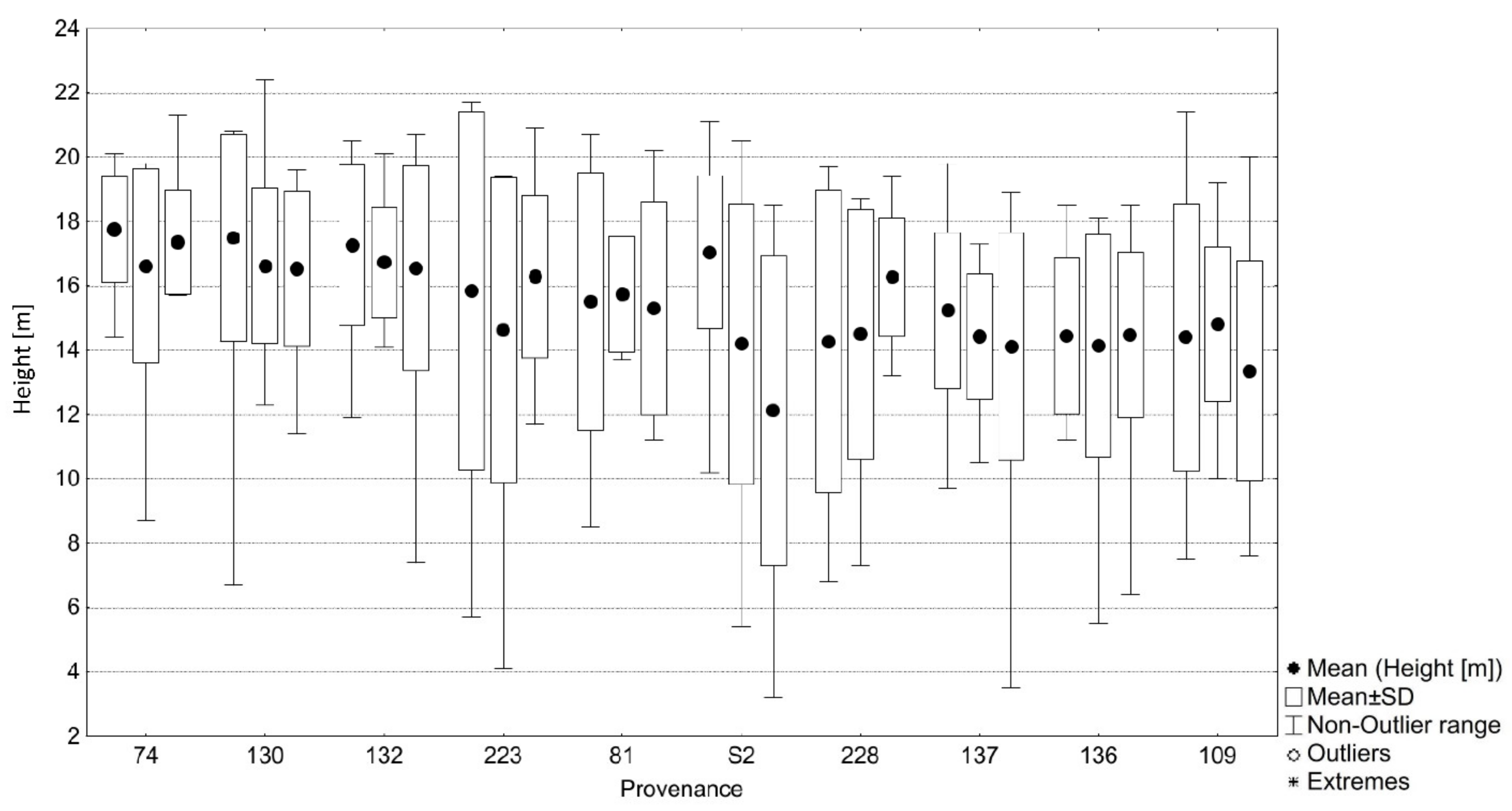
3.1. Comparison of Euro-Mediterranean Fir Species and Provenances in Trial 64, Písek
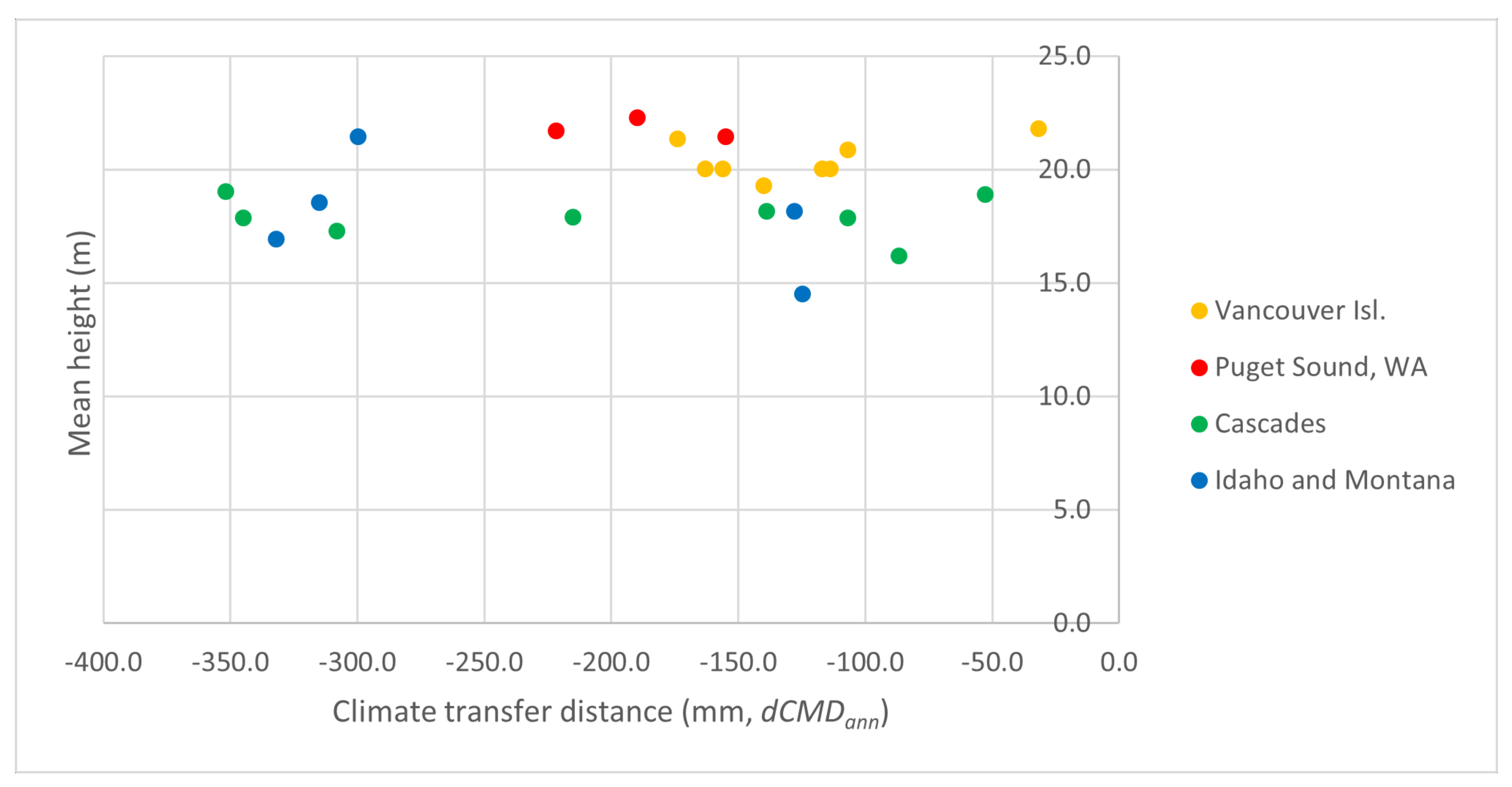
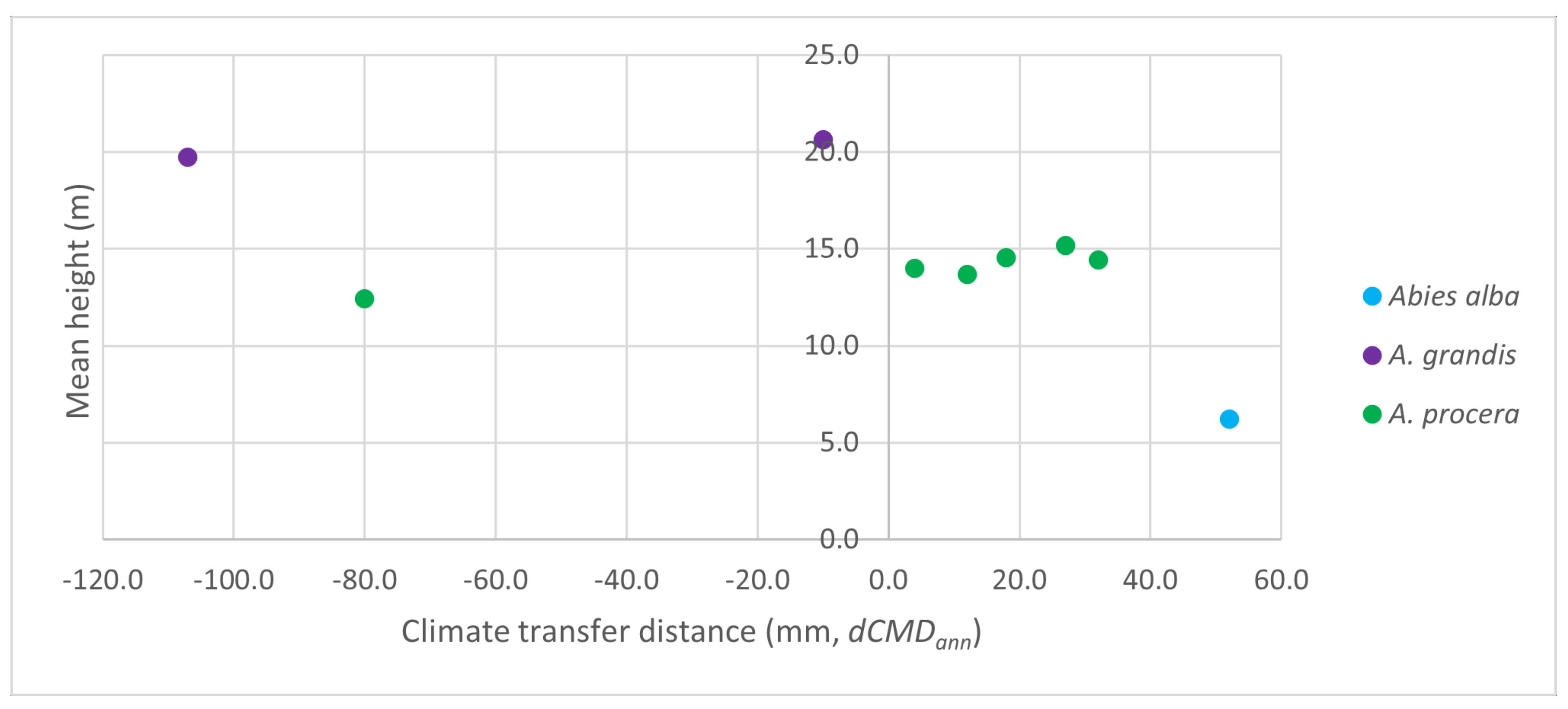
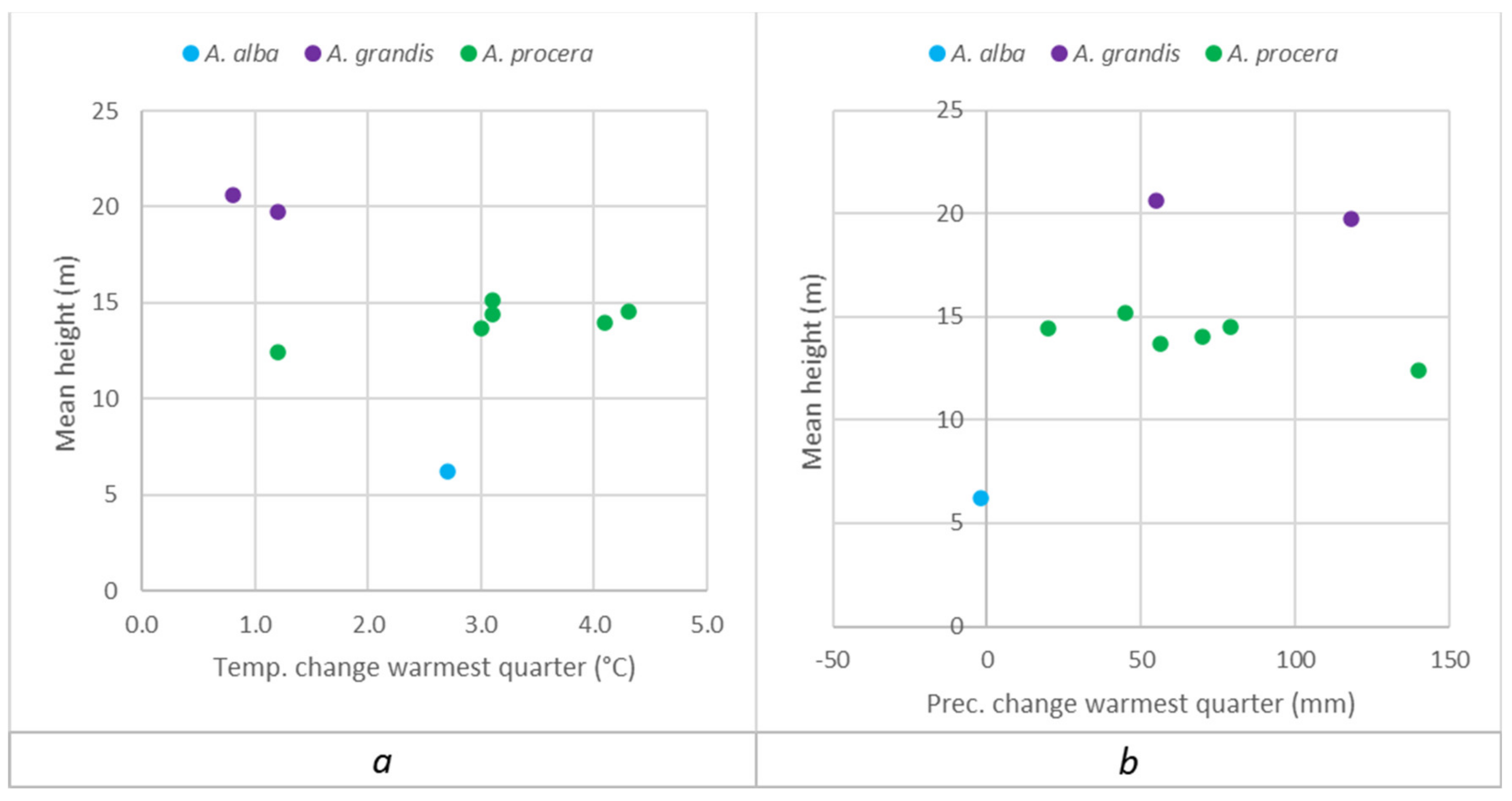
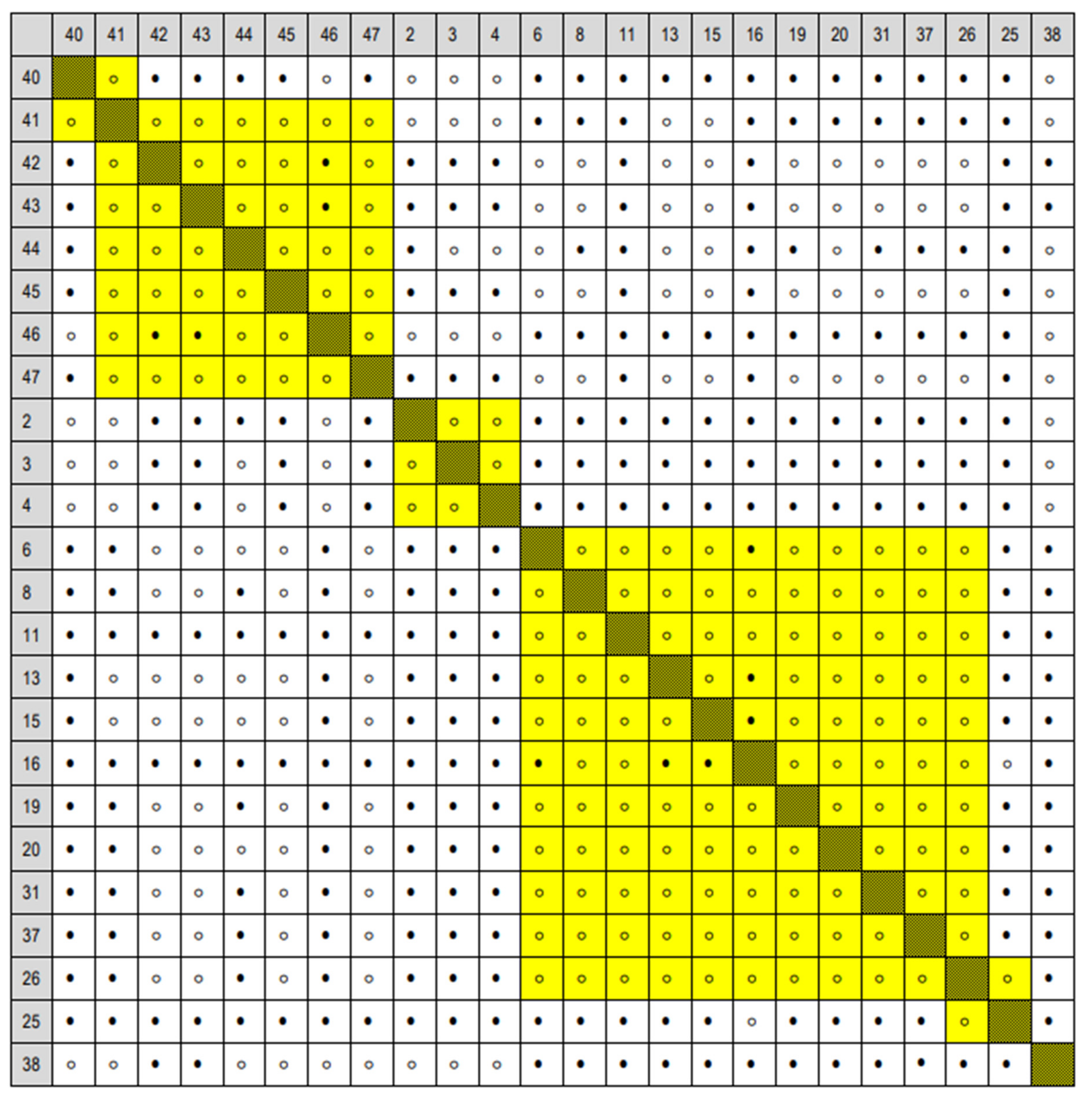
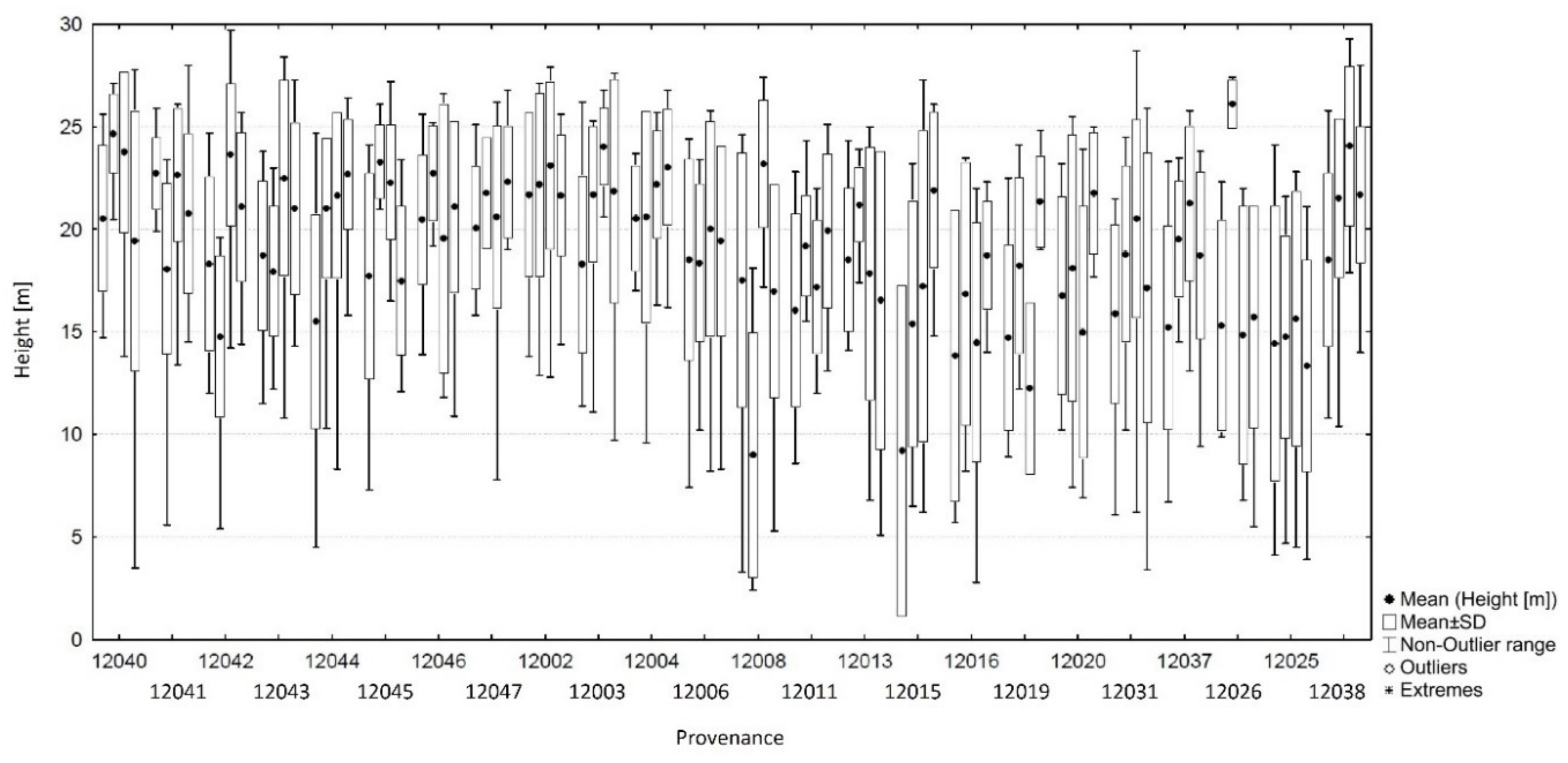
3.2. Comparison of Grand Fir Provenances in Trial 213, Zbiroh
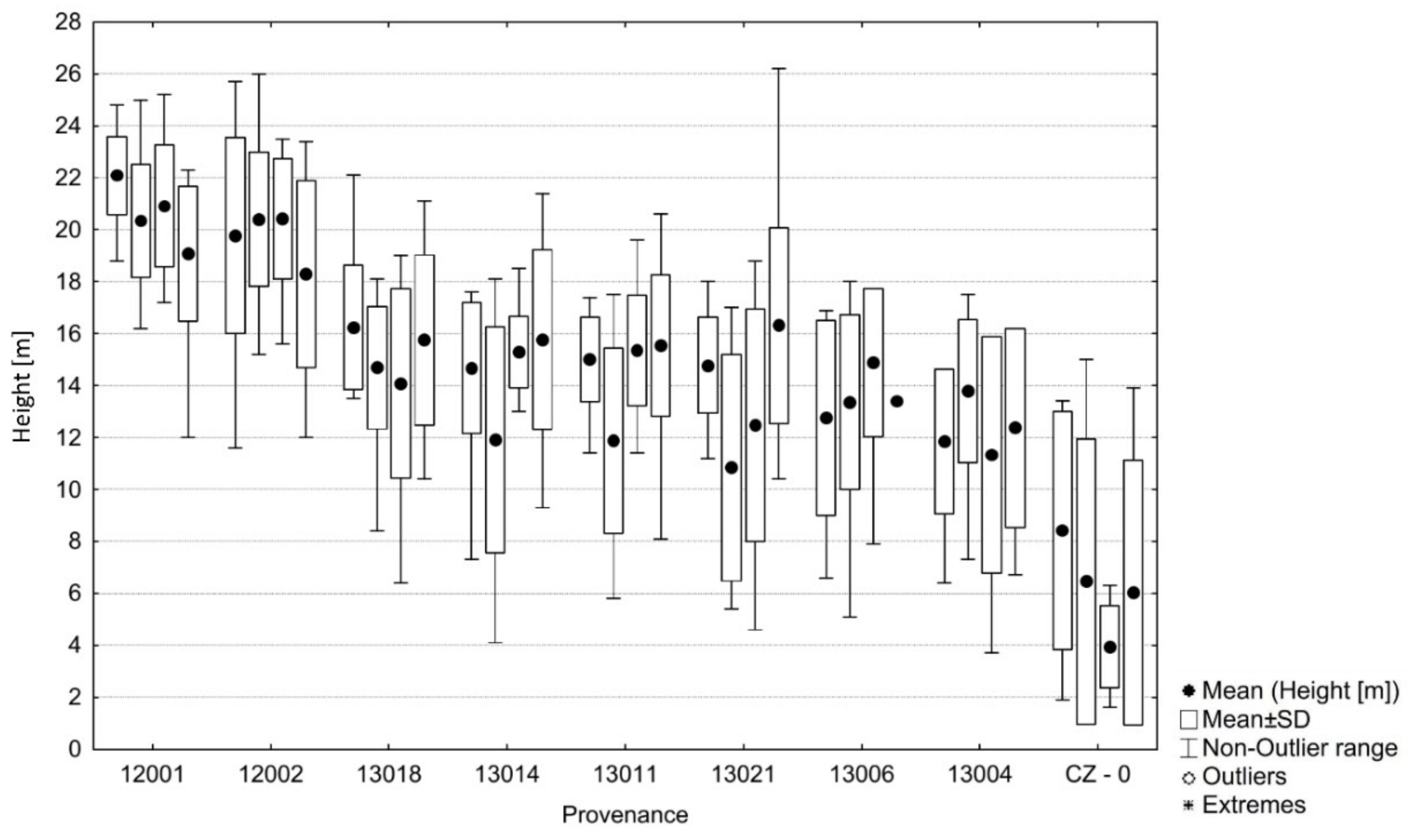
3.3. Comparison of Noble Fir Provenances in the Trial 219, Dražičky
4. Discussion
4.1. Adaptive Differentiation in Drought Tolerance
4.2. Projected Adaptive Response of Silver Fir to Future Climatic Challenges
4.3. Limitations of the Study Results
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
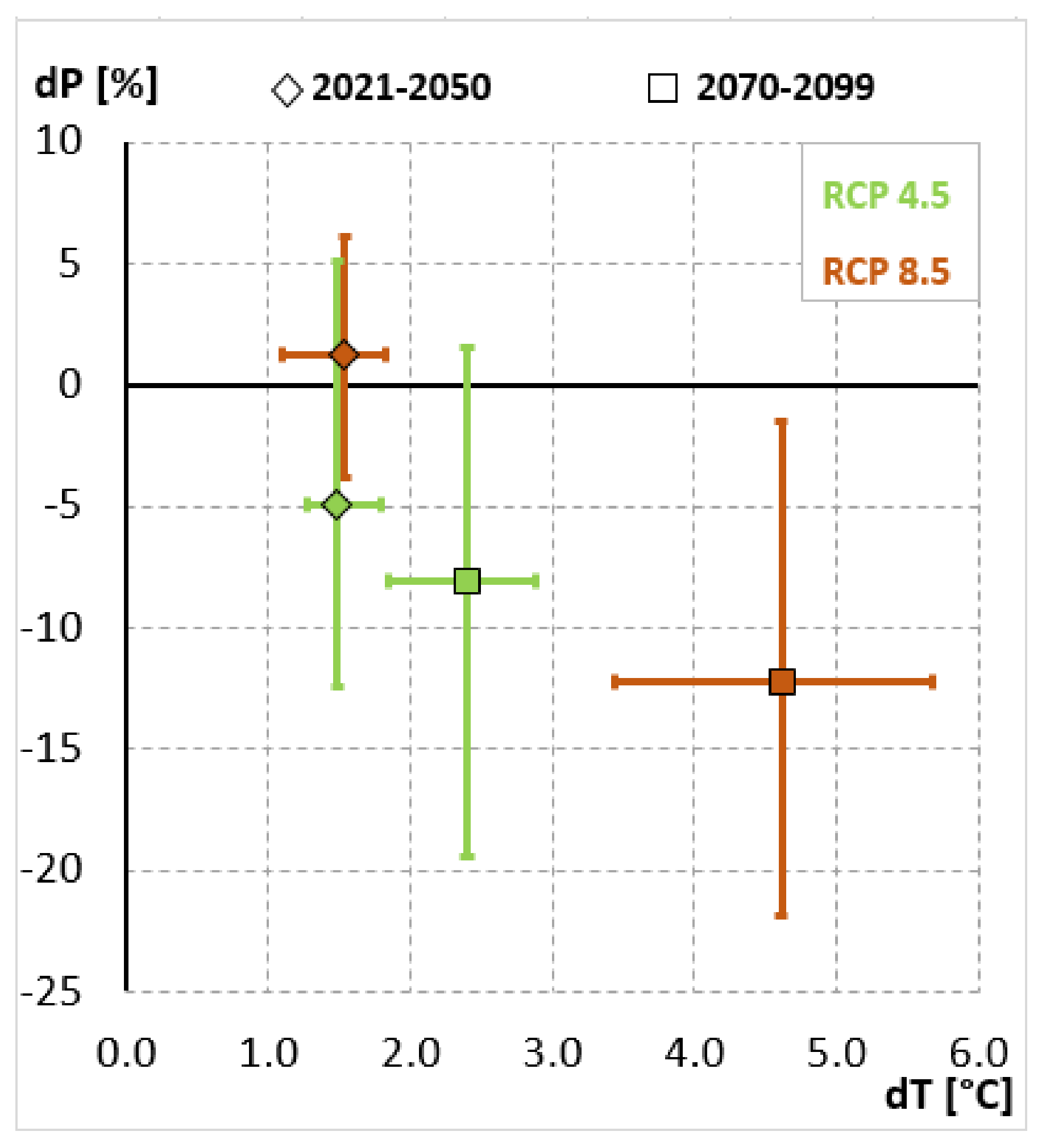
Appendix A.1. Additional Climate Data of Test Sites and Provenances

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Site Name | Meteorological Station Name | Altitude of Station | Distance from Site (km) | Altitude of Site (m a.s.l.) |
|---|---|---|---|---|
| Písek | Vráž | 433 | 19 | 395 |
| Zbiroh | Zbiroh | 480 | 11 | 450–460 |
| Dražičky | Tábor | 461 | 7 | 485 |
| Location | Climate Period | Mean Temperature | Mean Precipitation | CMD | |||
|---|---|---|---|---|---|---|---|
| Elevation (m) | (°C) | (mm) | (mm) | ||||
| Coordinates | Annual | Summer | Annual | Summer | Annual | Summer | |
| Písek. CZ Trial No.64 | 1911–1940 | 8.7 | 17.6 | 548 | 205 | 273 | 150 |
| 395 m | 1981–2010 * | 8.1 * | 17.2 * | 570 * | 229 * | 213 | 137 |
| Lat. 49.27 | 2040–2070 | 11.4 | 20.9 | 709 | 246 | 234 | 154 |
| Long. 14.33 | 2070–2100 | 13.2 | 23.2 | 691 | 222 | 315 | 211 |
| Zbiroh. CZ Trial No.213 | 1911–1940 | 7.7 | 16.4 | 533 | 191 | 234 | 134 |
| 456 m | 1981–2010* | 8.1 * | 17 * | 595 * | 241 * | 179 | 120 |
| Lat. 49.79 | 2040–2070 | 10.4 | 19.7 | 689 | 230 | 199 | 136 |
| Long. 13.64 | 2070–2100 | 12.1 | 22 | 673 | 209 | 274 | 189 |
| Dražičky. CZ Trial No. 219 | 1911–1940 | 8.1 | 16.9 | 539 | 206 | 250 | 137 |
| 488 m | 1981–2010 * | 8.1 * | 17.3 * | 592 * | 229 * | 189 | 122 |
| Lat. 49.39 | 2040–2070 | 10.9 | 20.3 | 696 | 247 | 209 | 140 |
| Long. 14.59 | 2070–2100 | 12.7 | 22.6 | 678 | 224 | 288 | 196 |
| Populations of the Provenance Trial 64 Písek, CZ | Past Climate at Origin (1911–1940) | Current Climate | Future Climate (2071–2100) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Nr. | Abies Species Name | Provenance Name | Alt. (m) | Ann. Mean Temp. (°C) | Ann. Mean Prec. (mm) | Ann. CMD (mm) | Mean Height (Year 2015) (m) | Temp. Annual Change (°C) | Prec. Annual Change (mm) | Ann. dCMD (mm) | Proj. Mean Height (m) *** |
| 74 | A. alba | Milevsko, Klučenice CZ | 410 | 8.4 | 492 | 284 | 17.19 | 4.8 | 199 | 31 | 16.17 |
| 81 | A. alba | Vyšší Brod, Vítkův Kámen CZ | 900 | 5.9 | 854 | 122 | 15.47 | 7.3 | −163 | 193 | 13.46 |
| 109 | A. cephalonica | Centr. Peloponnese, Vytina GR | 1250 | 10.9 | 794 | 573 | 14.19 | 2.3 | −103 | −258 | 14.94 |
| 130 | A. alba | Nasavrky, Podhůra CZ | 370 | 8.2 | 487 | 253 | 16.89 | 5.0 | 204 | 62 | 15.68 |
| 132 | A. alba | Rilskije gory, Borovec BG * | 1200 | 7.3 | 515 | 362 | 16.85 | 5.9 | 176 | −47 | 16.31 |
| 136 | A. cephalonica | Peloponnese, Vytina GR | 1010 | 12.2 | 753 | 626 | 14.34 | 1.0 | −62 | −311 | 15.41 |
| 137 | A. borisii-regis | Mt. Pindos, Pertuli GR * | 1200 | 9.6 | 727 | 503 | 14.59 | 3.6 | −36 | −188 | 14.91 |
| 223 | A. alba | Sanski Most BH * | 1050 | 6.9 | 943 | 170 | 15.61 | 6.3 | −252 | 145 | 13.89 |
| 228 | A. alba | Regello, Vallombrosa IT * | 1010 | 9.5 | 967 | 180 | 14.82 | 3.7 | −276 | 135 | 13.16 |
| S 2 | A. alba | Banská Bystrica, Radvaň SK * | 780 | 6.4 | 759 | 152 | 14.92 | 6.8 | −68 | 163 | 13.09 |
| Data and current climate of the test site ** | 395 | 8.1 | 570 | 213 | 15.49 | ||||||
| Future climate of the test site (2071–2100, RCP 8.5) | 395 | 13.2 | 691 | 315 | |||||||
| Populations of the Provenance Trial 64 Písek, CZ | Past Climate at Origin (1911–1940) | Current Climate | Future Climate (2071–2100) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Nr. | Abies Species Name | Provenance Name | Alt. (m) | Ann. Mean Temp. (°C) | Ann. Mean Prec. (mm) | Ann. CMD (mm) | Mean Height (Year 2015) (m) | Temp. Annual Change (°C) | Prec. Annual Change (mm) | Ann. dCMD (mm) | Proj. Mean Height (m) *** |
| 74 | A. alba | Milevsko, Klučenice CZ | 410 | 8.4 | 492 | 284 | 17.19 | 4.5 | 127 | 46 | 15.97 |
| 81 | A. alba | Vyšší Brod, Vítkův Kámen CZ | 900 | 5.9 | 854 | 122 | 15.47 | 4.5 | 226 | -39 | 17.13 |
| 109 | A. cephalonica | Centr. Peloponnese, Vytina GR | 1250 | 10.9 | 794 | 573 | 14.19 | 4.2 | −71 | 131 | 12.06 |
| 130 | A. alba | Nasavrky, Podhura CZ | 370 | 8.2 | 487 | 253 | 16.89 | 4.5 | 123 | 44 | 15.94 |
| 132 * | A. alba | Rilskije gory, Borovec BG | 1200 | 7.3 | 515 | 362 | 16.85 | 5.0 | 20 | 172 | 13.03 |
| 136 | A. cephalonica | Peloponnese, Vytina GR | 1010 | 12.2 | 753 | 626 | 14.34 | 4.2 | −65 | 135 | 12.77 |
| 137 * | A. borisii-regis | Mt. Pindos, Pertuli GR | 1200 | 9.6 | 727 | 503 | 14.59 | 4.8 | −31 | 162 | 11.26 |
| 223 * | A. alba | Sanski Most BH | 1050 | 6.9 | 943 | 170 | 15.61 | 4.7 | 130 | 97 | 14.78 |
| 228 * | A. alba | Regello, Vallombrosa IT | 1010 | 9.5 | 967 | 180 | 14.82 | 4.5 | 109 | 91 | 13.95 |
| S 2 * | A. alba | Banská Bystrica, Radvaň SK | 780 | 6.4 | 759 | 152 | 14.92 | 4.8 | 180 | 28 | 15.38 |
| Data and current climate of the test site ** | 395 | 8.1 | 570 | 213 | 15.49 | ||||||
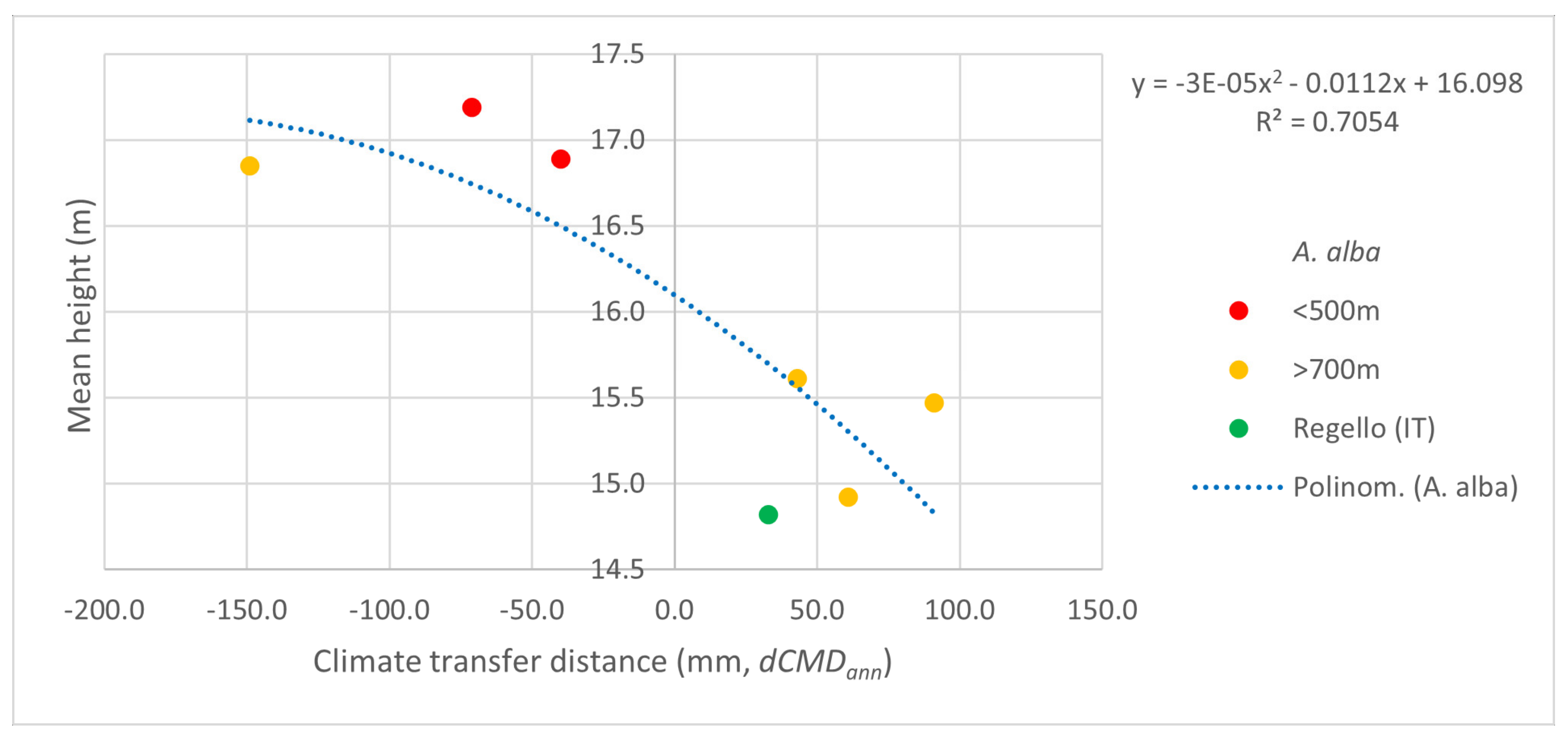
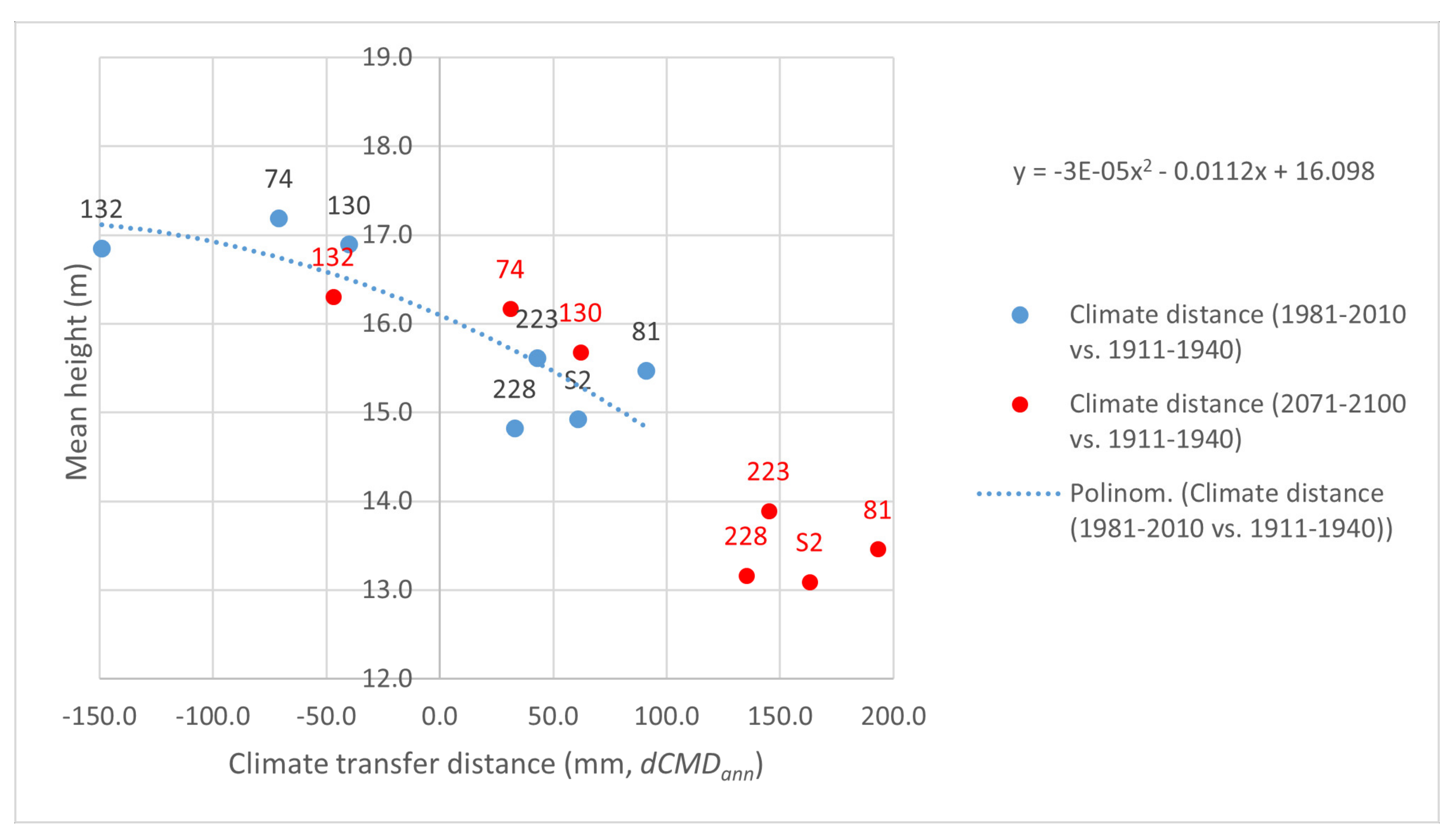
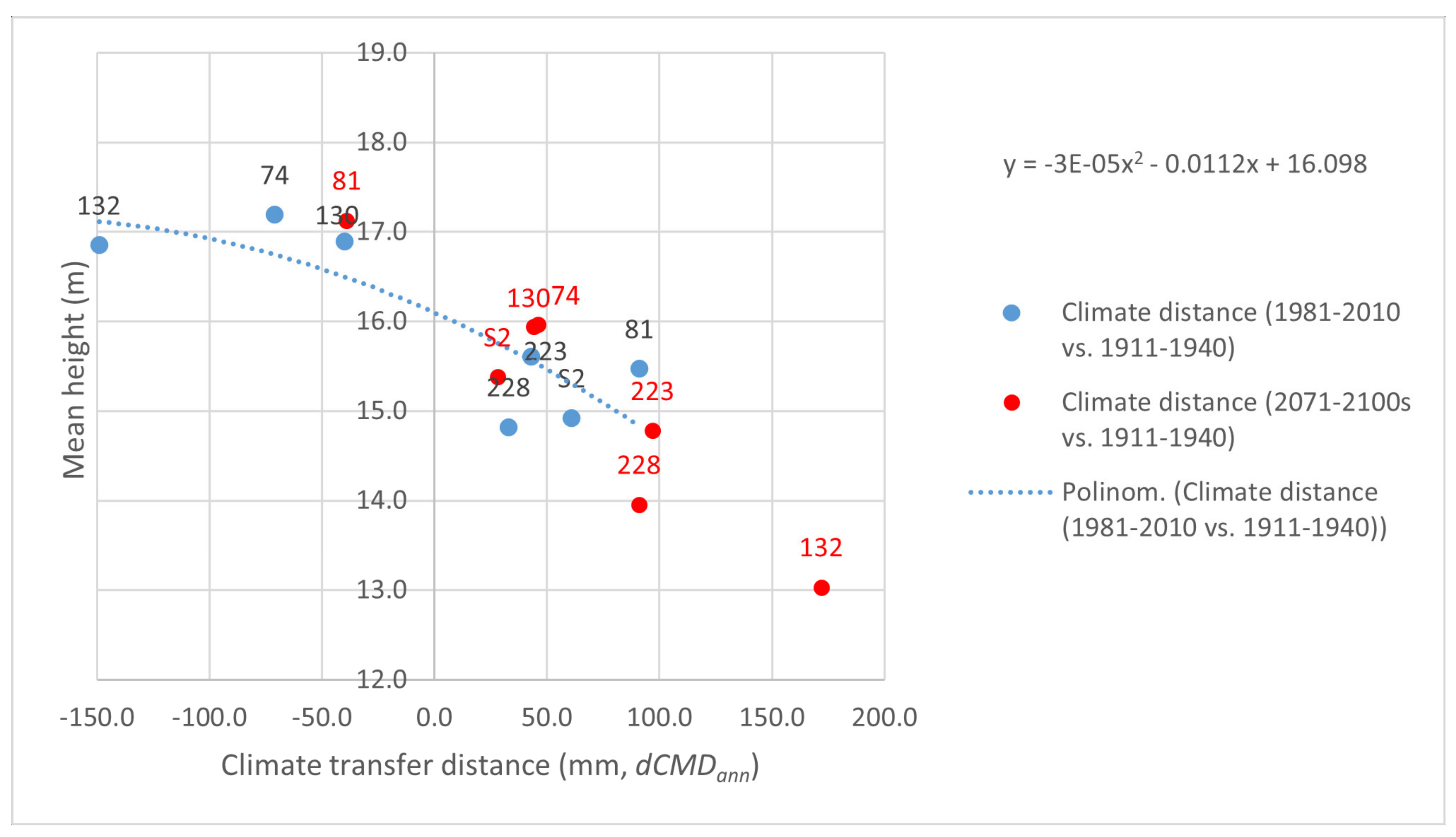
Appendix A.2. Separate Presentation of Height Responses to Temperature and Precipitation Changes in the Summer (Warmest) Quarter, Caused by Transfer of Populations to the Trials



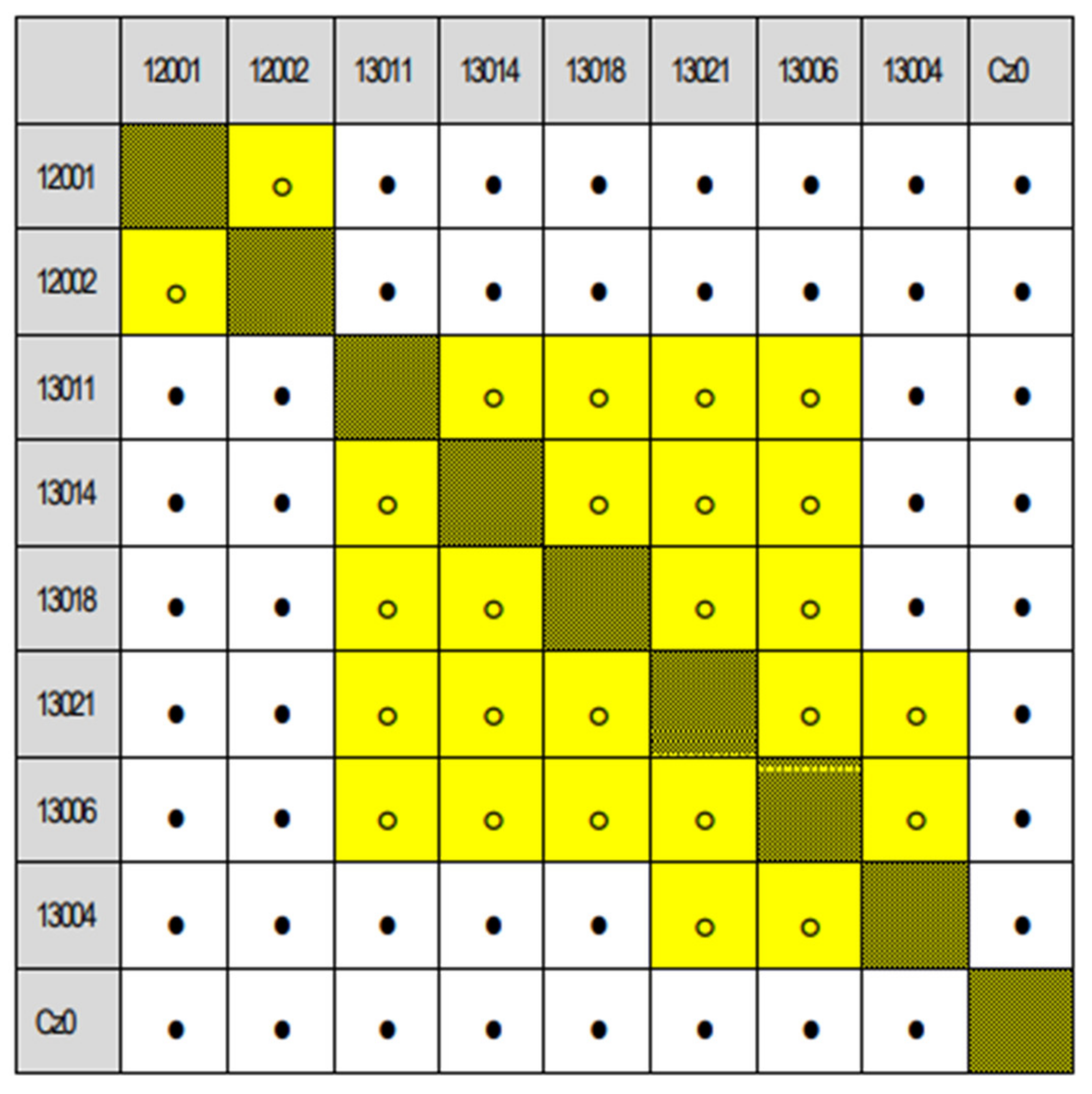
Appendix A.3. Matrices of Significance Calculated for the Three Provenance Tests, Based on Statistics in [20]



Appendix A.4. Variance of Populations Sorted by the Sequence of Significance Statistics (The Sequence of Provenances follow the Respective Matrices of Significance)



References
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 129. [Google Scholar] [CrossRef]
- Mátyás, C. Forecasts needed for retreating forests (Opinion). Nat. Intern. Weekl. J. Sci. 2010, 464, 1271. [Google Scholar]
- Mátyás, C.; Berki, I.; Bidló, A.; Csóka, G.; Czimber, K.; Führer, E.; Gálos, B.; Gribovszki, Z.; Illés, G.; Hirka, A.; et al. Sustainability of forest cover under climate change on the temperate-continental xeric limits. Forests 2018, 9, 489. [Google Scholar] [CrossRef] [Green Version]
- Spinoni, J.; Barbosa, P.; Bucchignani, E.; Cassano, J.; Cavazos, T.; Christensen, J.H.; Dosio, A. Future global meteorological drought hotspots: A study based on CORDEX data. J. Climate. 2019. [Google Scholar] [CrossRef]
- Gálos, B.; Führer, E.; Czimber, K.; Gulyás, K.; Bidló, A.; Hänsler, A.; Jacob, D.; Mátyás, C. Climatic threats determining future adaptive forest management—A case study of Zala County. Időjárás 2015, 119, 425–441. [Google Scholar]
- Czimber, K.; Gálos, B. A new decision support system to analyse the impacts of climate change on the Hungarian forestry and agricultural sectors. Scand. J. For. Res. 2016, 664–673. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Gutiérrez, E.; Popa, I.; Andreu-Hayles, L.; Motta, R.C.; Carrer, M. Distinct effects of climate warming on populations of silver fir (Abies alba) across Europe. J. Biogeogr. 2015, 42, 1150–1162. [Google Scholar] [CrossRef] [Green Version]
- Matías, L.; Gonzalez-Díaz, P.; Quero, J.L.; Camarero, J.J.; Lloret, F.; Jump, A.S. Role of geographical provenance in the response of silver fir seedlings to experimental warming and drought. Tree Physiol. 2016, 36, 1236–1246. [Google Scholar] [CrossRef] [Green Version]
- Brázdil, R.; Trnka, M. Sucho v českých zemích: Minulost, současnost, budoucnost. In Drought in the Czech Lands: Past, Present, Future, 1st ed.; Research Centre for Global Change, Academy of Science of the Czech Republic: Brno, Czech Republic; UNI Press: Turnov, Czech Republic, 2015. (In Czech) [Google Scholar]
- Šindelář, J.; Frýdl, J. K problematice genetických změn v populacích lesních dřevin v důsledku narušování lesů škodlivými vlivy prostředí. Problems of genetic changes in forest tree populations due to disturbances by harmful environmental influences. Zprávy Lesn. Výzk. 2008, 53, 116–119. (In Czech) [Google Scholar]
- Lebourgeois, F.; Rathgeber, C.B.K.; Ulrich, E. Sensitivity of French temperate coniferous forests to climate variability and extreme events (Abies alba, Picea abies and Pinus sylvestris). J. Veget. Sci. 2010, 21, 364–376. [Google Scholar] [CrossRef]
- George, J.-P.; Grabner, M.; Campelo, F.; Karanitsch-Ackerl, S.; Mayer, K.; Klumpp, R.T.; Schüler, S. Intra-specific variation in growth and wood density traits under water-limited conditions: Long-term-, short-term-, and sudden responses of four conifer tree species. Sci. Total Environ. 2019, 660, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Vitasse, Y.; Bottero, A.; Rebetez, M.; Conedera, M.; Augustin, S.; Brang, P.; Tinner, W. What is the potential of silver fir to thrive under warmer and drier climate? Eur. J. Forest Res. 2019. [Google Scholar] [CrossRef]
- Benito-Garzón, M.; Robson, T.M.; Hampe, A. ΔTraitSDM: Species distribution models that account for local adaptation and phenotypic plasticity. New Phytol. 2019, 222, 1757–1765. [Google Scholar] [CrossRef] [PubMed]
- Bouriaud, O.; Popa, I. Comparative dendroclimatic study of Scots pine, Norway spruce, and silver fir in the Vrancea Range, Eastern Carpathian Mountains. Trees 2008, 23, 95–106. [Google Scholar] [CrossRef]
- Tinner, W.; Colombaroli, D.; Heiri, O.; Henne, P.D.; Steinacher, M.; Untenecker, J.; Conedera, M. The past ecology of Abies alba provides new perspectives on future responses of silver fir forests to global warming. Ecol. Monogr. 2013, 83, 419–439. [Google Scholar] [CrossRef] [Green Version]
- Zang, C.; Hartl-Meier, C.; Dittmar, C.; Rothe, A.; Menzel, A. Patterns of drought tolerance in major European temperate forest trees: Climatic drivers and levels of variability. Global Chang. Biol. 2014, 20, 3767–3779. [Google Scholar] [CrossRef]
- Vitali, V.; Büntgen, U.; Bauhus, J. Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in south-western Germany. Glob. Chang. Biol. 2017, 23, 5108–5119. [Google Scholar] [CrossRef] [PubMed]
- Kapeller, S.; Schüler, S.; Huber, G.; Božič, G.; Wohlgemuth, T.; Klumpp, R. Provenance Trials in Alpine Range—Review and Perspectives for Applications in Climate Change. In Management Strategies to Adapt Alpine Space Forests to Climate Change Risks; Cerbu, G., Ed.; IntechOpen: London, UK, 2013; ISBN 978-953-51-1194-8. [Google Scholar] [CrossRef] [Green Version]
- Frýdl, J.; Dostál, J.; Beran, F.; Čáp, J.; Fulín, M.; Frampton, J.; Božič, G.; Mátyás, C. Exotic Abies Species in Czech Provenance Trials: Assessment after Four Decades. Acta Silv. Lign. Hung. 2018, 14, 1, 9–34. [Google Scholar] [CrossRef]
- Sáenz-Romero, C.; Mendoza-Maya, E.; Gómez-Pineda, E.; Blanco-García, A.; Endara-Agramont, A.R.; Lindig-Cisneros, R.; Vargas-Hernández, J.J. Recent evidence of Mexican temperate forest decline and the need for ex situ conservation, assisted migration, and translocation of species ensembles as adaptive management to face projected climatic change impacts in a megadiverse country. Can. J. For. Res. 2020, 50, 843–854. [Google Scholar] [CrossRef]
- Booth, T.H. Assessing species climatic requirements beyond the realized niche: Some lessons mainly from tree species distribution modelling. Clim. Chang. 2017, 145, 259–271. [Google Scholar] [CrossRef]
- Mátyás, C.; Yeatman, C.W. Effect of geographical transfer on growth and survival of jack pine (Pinus banksiana Lamb.) populations. Silvae Genet. 1992, 41, 370–376. [Google Scholar]
- Mátyás, C. Modelling climate-change effects with provenance test data. Tree Physiol. 1994, 14, 797–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagnard, F.; Barberot, C.; Fady, B. Structure of genetic diversity in Abies alba Mill. from southwestern Alps: Multivariate analysis of adaptive and non-adaptive traits for conservation in France. For. Ecol. Manag. 2002, 157, 175–189. [Google Scholar] [CrossRef]
- Latreille, A.C.; Pichot, C. Local-scale diversity and adaptation along elevational gradients assessed by reciprocal transplant experiments: Lack of local adaptation in silver fir populations. Ann. For. Sci. 2017, 74, 77. [Google Scholar] [CrossRef] [Green Version]
- Mayer, H.; Reimoser, F.; Kral, F. Ergebnisse des Internationalen Tannenherkunftsversuches Wien 1967—1978. Morphologie und Wuchsverhalten der Provenienzen. Cbl. Ges. Forstw. 1982, 99, 169–191. [Google Scholar]
- Ruetz, W.F.; Dimpflmeier, R.; Kleinschmit, J.; Svolba, J.; Weisgerber, H.; Rau, H.M. The IUFRO Abies procera provenance trial in the Federal Republic of Germany: Field results at age 9 and 10 years. In Proceeding Joint Meeting Western Forest Genetics Assoc. and IUFRO Working Parties S2.02-05, 06, 12 and 14; Weyerhaeuser Co.: Olympia, WA, USA, 1990. [Google Scholar]
- Ruetz, W.; Stimm, B. Der Süddeutsche Weißtannen-Provenienzversuch: IV. Entwicklung der Herkünfte der Aussaat 1982 auf den Versuchsflächen in Bayern bis zum Alter von 12 Jahren. In Ergebnisse des 7. Weisstannen-Symposiums in Altensteig, Deutschland; Eder, W., Ed.; IUFR0-Verlag: Mainz, Germany, 1994; pp. 17–29. [Google Scholar]
- Klumpp, R.T. Der Tannenprovenienzversuch “Knödelhütte Wien 1967”: Ergebnisse im Alter 24. Mitt. Forsch. Anst. Waldökol. Forstw. RheinIand-Pfalz. 2002, 50/03, 44–49. [Google Scholar]
- Gugerli, F.; Frank, A.; Rellstab, C.; Pluess, A.R.; Moser, B.; Ahrend, M.; Sperisen, C.; Wohlgemuth, T.; Heiri, C. Genetische Variation und lokale Anpassung bei Waldbaumarten im Zeichen des Klimawandels. (Genetic variation and local Adaptation in forest tree species under climate change.). In Wald im Klimawandel. Grundlagen für Adaptationsstrategien. (Forests in Climate Change. Basics of Adaptation Strategies.); Pluess, A.R., Augustin, S., Brang, P., Eds.; BAFU, WSL Birmensdorf, Haupt Verl.: Bern, Switzerland, 2016; pp. 93–118. [Google Scholar]
- Frank, A.; Sperisen, C.; Howe, G.T.; Brang, P.; Walthert, L.; St. Clair, J.B.; Heiri, C. Distinct genecological patterns in seedlings of Norway spruce and silver fir from a mountainous landscape. Ecology 2017, 98, 211–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csilléry, K.; Buchmann, N.; Fady, B. Adaptation to drought is coupled with slow growth, but independent from phenology in marginal silver fir (Abies alba Mill.) populations. Evol. Appl. 2020, 13. [Google Scholar] [CrossRef]
- Hansen, J.K.; Larsen, J.B. European silver fir (Abies alba Mill.) provenances from Calabria, Southern Italy: 15-year results from Danish provenance field trials. Eur. J. For. Res. 2004, 123, 127–138. [Google Scholar] [CrossRef]
- George, J.P.; Schueler, S.; Karanitsch-Ackerl, S.; Mayer, K.; Klumpp, R.T.; Grabner, M. Inter- and intra-specific variation in drought sensitivity in Abies spec. and its relation to wood density and growth traits. Agric. For. Meteorol. 2015, 214–215, 430–443. [Google Scholar] [CrossRef] [Green Version]
- Bošel’a, M. Jedl’a biela: Perspektívna drevina v čase meniacej sa klímy? [Silver fir: A species with perspective in case if climate changes?]. In Zachovanie druhovej rozmanitosti lesov Slovenska v čase globálnych environmentálnych zmien. [Conservation of species diversity of Slovakia’s forests in times of global environmental change]; Mális, F., Ed.; Bulletin Správy; Forestry Faculty, TU Zvolen: Zvolen, Slovakia, 2017; pp. 3–9. (In Slovak) [Google Scholar]
- Csilléry, K.; Ovaskainen, O.; Sperisen, C.; Buchmann, N.; Widmer, A.; Gugerli, F. Adaptation to local climate in multi-trait space: Evidence from silver fir (Abies alba Mill.) populations across a heterogeneous environment. Heredity 2020, 124, 77–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šindelář, J.; Beran, F. Comparison of some exotic species of Abies genus with chosen silver fir provenances on the plots of town Písek. Comm. Inst. For. Bohem. 2008, 24, 99–113. [Google Scholar]
- Šindelář, J.; Beran, F. Exotic species of fir (Abies spec. div.) at the age of 30 years in the nature forest region No. 10-Středočeská pahorkatina (Central Bohemian Upland). Comm. Inst. For. Bohem. 2008, 24, 115–130. [Google Scholar]
- Mátyás, C. Adaptive pattern of phenotypic plasticity and inherent growth reveal the potential for assisted transfer in sessile oak (Quercus petraea L.). For. Ecol. Manag. 2021, 482, 118832. [Google Scholar] [CrossRef]
- Nielsen, R. The Little Green Handbook; Scribe Publications: London, UK, 2006; ISBN 0312425813. Available online: http://home.iprimus.com.au/nielsens/ (accessed on 2 December 2020).
- Steffen, W.; Broadgate, W.; Deutsch, L.; Gaffney, O.; Ludwig, C. The Trajectory of the Anthropocene: The Great Acceleration. Anthr. Rev. 2015, 2, 81–98. Available online: http://hdl.handle.net/1885/66463 (accessed on 4 December 2020). [CrossRef]
- Marchi, M.; Castellanos-acuña, D.; Hamann, A.; Wang, T.; Ray, D.; Menzel, A. ClimateEU, scale-free climate normals, historical time series, and future projections for Europe. Sci. Data 2020, 7, 428. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Hamann, A.; Spittlehouse, D.L.; Murdock, T.Q. ClimateWNA-high-resolution spatial climate data for western North America. J. Appl. Meteorol. Climatol. 2012, 51, 16–29. [Google Scholar] [CrossRef] [Green Version]
- Hargreaves, G.H. Defining and using reference evapotranspiration. J. Irrigat. Drain. Eng. 1994, 120, 6. [Google Scholar] [CrossRef]
- QC-Expert 3.1 TriloByte Statistical Software. TriloByte Ltd., Pardubice—Staré Hradiště, Czech Republic. Available online: http://www.trilobyte.cz/Nase-Produkty/QC-Expert.html(accessed on 10 November 2020).
- CEDA Archive. Climate Research Unit, UK, Time Series Version 3.23, High Resolution Gridded Data. 2018. Available online: http://catalogue.ceda.ac.uk/uuid/5dca9487dc614711a3a933e44a933ad3 (accessed on 10 November 2020).
- Alizoti, P.; Bastien, J.C.; Chakraborty, D.; Klisz, M.; Kroon, J.; Neophytou, C.; Schueler, S.; van Loo, M.; Westergren, M.; Konnert, M.; et al. Non-native forest tree species in Europe: The question of seed origin in afforestation. Forestry 2019. Available online: http://mc.manuscriptcentral.com/forest (accessed on 10 November 2020).
- Mátyás, C.; Bozic, G.; Gömöry, D.; Ivankovic, M.; Rasztovits, E. Juvenile growth response of European beech (Fagus sylvatica L.) to sudden change of climatic environment in SE European trials. iForest, J. Biogeosc. For. 2009, 2, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Konôpková, A.; Pšidová, E.; Kurjak, D.; Stojnić, S.; Petrík, C.P.; Fleischer, A.P.; Ditmarová, L. Photosynthetic performance of silver fir (Abies alba) of different origins under suboptimal growing conditions. Funct. Plant. Biol. 2020, 47. [Google Scholar] [CrossRef]
- Mosca, E.; Cruz, F.; Gómez-Garrido, J.; Bianco, L.; Rellstab, C.; Brodbeck, S.; Neale, D.B. A Reference Genome Sequence for the European Silver Fir (Abies alba Mill.): A Community-Generated Genomic Resource. G3 Genes Genomes Genetics 2019, 9, 2039–2049. [Google Scholar] [CrossRef] [Green Version]
- Fulín, M.; Remeš, J.; Tauchman, P. Růst a produkce jedle obrovské (Abies grandis Lindl.) ve srovnání s jinými jehličnany. Growth and production of grand fir (Abies grandis Lindl.) compared with other conifer species. Zprávy lesn. výzk. 2013, 58, 186–192. (in Czech). [Google Scholar]
- Roschanski, A.M.; Csilléry, K.; Liepelt, S.; Oddou-Muratorio, S.; Ziegenhagen, B.; Huard, F.; Fady, B. Evidence of divergent selection for drought and cold tolerance at landscape and local scales in Abies alba Mill. in the French Mediterranean Alps. Mol. Ecol. 2016, 25, 776–794. [Google Scholar] [CrossRef] [PubMed]
- Čáp, J.; Novotný, P.; Frýdl, J. Vyhodnocení provenienční výzkumné plochy s jedlí bělokorou (Abies alba Mill.) č. 57-Lesy Jíloviště, Cukrák ve věku 35 let. Evaluation of a provenance research trial with silver fir at the age of 35 years. Zprávy Lesn. Výzk. 2009, 54, 33–43. (In Czech) [Google Scholar]
- Rehfeldt, G.E.; Leites, L.P.; Joyce, D.G.; Weiskittel, A.R. Role of population genetics in guiding ecological responses to climate. Glob. Chang. Biol. 2018, 24, 858–868. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Warwell, M.V.; Monserud, R.A. Species, climatypes, climate change and forest health: A conversion of science to practice for inland northwest (USA) forests. Forests 2020, 11, 1237. [Google Scholar] [CrossRef]
- Mihai, G.; Alexandru, A.M.; Stoica, E.; Birsan, M.V. Intraspecific growth response to drought of Abies alba in the Southeastern Carpathians. Forests 2021, 12, 387. [Google Scholar] [CrossRef]
- Wilczyński, S.; Kulej, M. The growth of seven Abies grandis provenances in the climatic conditions of the Polish Carpathian Mountains. Dendrobiology 2019, 81, 1–13. [Google Scholar] [CrossRef]
- Grady, K.C.; Kolb, T.E.; Ikeda, D.H.; Whitham, T.G. A bridge too far: Cold and pathogen constraints to assisted migration of riparian forests. Restor. Ecol. 2015, 23, 811–820. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/rec.12245 (accessed on 5 December 2020). [CrossRef]
- Adams, H.D.; Barron-Gafford, G.A.; Minor, R.L.; Gardea, A.A.; Bentley, L.P.; Law, D.J.; Huxman, T.E. Temperature response surfaces for mortality risk of tree species with future drought. Environ. Res. Lett. 2017, 12, 115014. [Google Scholar] [CrossRef]
- Berki, I.; Rasztovits, E.; Móricz, N.; Kolozs, L. The role of tree mortality in vitality assessment of sessile oak forests. South-East Eur. For. 2016, 7, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Střelcová, K. Stres suchom a lesné porasty. [Drought stress and forest trees]. In Zachovanie Druhovej Rozmanitosti Lesov Slovenska v čase Globálnych Environmentálnych Zmien. [Conservation of species diversity of Slovakia’s forests in times of global environmental change]; Mális, F., Ed.; Bulletin Správy; Forestry Faculty, TU Zvolen: Zvolen, Slovakia, 2017; pp. 59–71. (In Slovak) [Google Scholar]










| Trial Name | Year of Planting | Altitude (m) | Longitude (Decimal) | Latitude (Decimal) | Age at Measurement (years) | Trial Mean Height (m) | Mean Tree Density (% from Planted) |
|---|---|---|---|---|---|---|---|
| Písek | 1976 | 395 | 14.33° E | 49.27° N | 40 | 15.49 | 28.5 a |
| Zbiroh | 1980 | 450–460 | 13.64° E | 49.79° N | 36 | 19.26 | 50.1 |
| Dražičky | 1984 | 485 | 14.59° E | 49.39° N | 32 | 14.54 | 55.9 |
| Trial Name | Annual Mean Temp. (°C) | Mean Temp. in Warmest Quarter (°C) | Annual Mean Precip. (mm) | Mean Precip. in Warmest Quarter (mm) | Mean Annual Moisture Deficit (CMDann) |
|---|---|---|---|---|---|
| Písek | 8.1 | 17.2 | 570 | 229 | 213 |
| Zbiroh | 8.1 | 17.0 | 595 | 241 | 179 |
| Dražičky | 8.1 | 17.3 | 592 | 229 | 189 |
| Populations of the Provenance Trial 64 Písek, CZ | Past Climate at Origin (1911–1940) | Climate Distance (D) | Data 2015 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Prov No. | Abies Species Name | Provenance Name | Alt. (m) | Long. E | Lat. N | January Mean Temp. (°C) | Annual Mean Temp. (°C) | Mean Temp. Warmest Quarter (°C) | Annual Mean Prec. (mm) | Mean Prec. Warmest Quarter (mm) | CMD (mm, Ann.) | Temp. Change Warmest Quarter (°C) | Prec. Change Warmest Quarter (mm) | dCMD (Index Change, mm) | Mean Height (m) | Mean Tree Density (% From Planted) |
| 74 | A. alba | Milevsko, Klučenice CZ | 410 | 14.2° | 49.6° | −2 | 8.4 | 17.3 | 492 | 189 | 284 | −0.1 | 40.0 | −71 | 17.19 | 22.0 |
| 81 | A. alba | Vyšší Brod, Vítkův Kámen CZ | 900 | 14.3° | 48.6° | −4.2 | 5.9 | 14.5 | 854 | 296 | 122 | 2.7 | −67.0 | 91 | 15.47 | 10.0 |
| 89 * | A. cilicica | Kammouha LIB | 1100 | 36.0° | 34.0° | 6.3 | 14.9 | 22.2 | 1167 | 4 | 763 | −5.0 | 225.0 | −550 | 0.00 | 0.0 |
| 109 | A. cephalonica | Centr. Peloponnese, Vytina GR | 1250 | 22.1° | 37.6° | 1.9 | 10.9 | 18.7 | 794 | 51 | 573 | −1.5 | 178.0 | −360 | 14.19 | 29.3 |
| 121 * | A. cilicica | Djebel el Chouk, Lattakia SYR | 1300 | 36.0° | 35.8° | 2.2 | 12.6 | 20 | 1039 | 24 | 525 | −2.8 | 205.0 | −312 | 0.00 | 0.0 |
| 130 | A. alba | Nasavrky, Podhůra CZ | 370 | 15.8° | 49.8° | −2.7 | 8.2 | 16.8 | 487 | 196 | 253 | 0.4 | 33.0 | −40 | 16.89 | 32.7 |
| 132 * | A. alba | Rilskije gory, Borovec BG | 1200 | 23.6° | 42.2° | −3 | 7.3 | 15.7 | 515 | 137 | 362 | 1.5 | 92.0 | −149 | 16.85 | 31.3 |
| 135 * | A. pinsapo | Malaga, La Yunquera ES | 878 | 4.7° W | 36.7° | 6.8 | 14.7 | 22.2 | 633 | 18 | 711 | −5.0 | 211.0 | −498 | 0.00 | 0.0 |
| 136 | A. cephalonica | Peloponnese, Vytina GR | 1010 | 22.2° | 37.7° | 3.4 | 12.2 | 20 | 753 | 48 | 626 | −2.8 | 181.0 | −413 | 14.34 | 34.7 |
| 137 * | A. borisii-regis | Mt. Pindos, Pertuli GR | 1200 | 21.3° | 39.8° | −0.3 | 9.6 | 18.2 | 727 | 88 | 503 | −1.0 | 141.0 | −290 | 14.59 | 35.3 |
| 223 * | A. alba | Sanski Most BH | 1050 | 16.6° | 44.6° | −3.5 | 6.9 | 15.3 | 943 | 226 | 170 | 1.9 | 3.0 | 43 | 15.61 | 28.0 |
| 228 * | A. alba | Regello, Vallombrosa IT | 1010 | 11.5° | 43.7° | 0.2 | 9.5 | 17.5 | 967 | 163 | 180 | −0.3 | 66.0 | 33 | 14.82 | 30.7 |
| S 2 * | A. alba | Banská Bystrica, Radvaň SK | 780 | 19.0° | 48.7° | −5.1 | 6.4 | 15.1 | 759 | 258 | 152 | 2.1 | −29.0 | 61 | 14.92 | 31.3 |
| Data and current climate of the test site ** | 395 | 14.3° | 49.2° | −1.5 | 8.1 | 17.2 | 570 | 229 | 213 | 15.49 | 28.5 | |||||
| Populations of the Provenance Trial 213 Zbiroh, CZ | Past Climate at Origin (1911–1940) | Climate Distance (D) | Data 2015 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Prov No. | Abies Species Name | Provenance Name | Alt. (m) | Long. W | Lat. N | January Mean Temp. (°C) | Annual Mean Temp. (°C) | Mean Temp. Warmest Quarter (°C) | Annual Mean Prec. (mm) | Mean Prec. Warmest Quarter (mm) | CMD (mm, ann.) | Temp. Change Warmest Quarter (°C) | Prec. Change Warmest Quarter (mm) | dCMD (Index Change, mm) | Mean Height (m) | Mean Tree Density (% from Planted) |
| 12040 | A. grandis | Salmon River CAN/BC * | 50 | 125.8° | 50.3° | 0.6 | 8.4 | 15.6 | 1444 | 150 | 211 | 1.4 | 91.0 | −32 | 21.81 | 44.0 |
| 12041 | A. grandis | Oyster Bay CAN/BC | 5 | 125.2° | 49.9° | 1.2 | 9.1 | 16.3 | 1137 | 121 | 286 | 0.7 | 120.0 | −107 | 20.90 | 70.0 |
| 12042 | A. grandis | Buckley Bay CAN/BC | 45 | 124.9° | 49.5° | 0 | 8.8 | 16.3 | 1391 | 87 | 319 | 0.7 | 154.0 | −140 | 19.32 | 62.0 |
| 12043 | A. grandis | Sproat Lake CAN/BC | 25 | 125.0° | 49.3° | 0.8 | 9.4 | 17 | 1827 | 117 | 293 | 0.0 | 124.0 | −114 | 20.05 | 58.0 |
| 12044 | A. grandis | Kay Road CAN/BC | 50 | 124.3° | 49.3° | 2 | 9.4 | 16.5 | 863 | 85 | 342 | 0.5 | 156.0 | −163 | 20.02 | 72.0 |
| 12045 | A. grandis | Yellow Point CAN/BC | 30 | 123.8° | 49.1° | 2.7 | 9.7 | 16.6 | 916 | 81 | 335 | 0.4 | 160.0 | −156 | 20.03 | 57.0 |
| 12046 | A. grandis | Mount Provost CAN/BC | 75 | 123.8° | 48.8° | 1.8 | 9.5 | 16.7 | 1199 | 78 | 353 | 0.3 | 163.0 | −174 | 21.38 | 63.0 |
| 12047 | A. grandis | Sooke CAN/BC | 20 | 123.8° | 48.4° | 2.9 | 9.6 | 14.9 | 1111 | 72 | 296 | 2.1 | 169.0 | −117 | 20.05 | 60.0 |
| 12002 | A. grandis | Tulalip USA/WA | 30 | 122.3° | 48.1° | 3.4 | 9.5 | 14.8 | 651 | 53 | 369 | 2.2 | 188.0 | −190 | 22.29 | 48.0 |
| 12003 | A. grandis | Indian Creek USA/WA | 140 | 123.6° | 48.1° | 2.1 | 9 | 14.7 | 1011 | 64 | 334 | 2.3 | 177.0 | −155 | 21.48 | 57.0 |
| 12004 | A. grandis | Gardiner USA/WA | 30 | 122.9° | 48.1° | 2.9 | 9.6 | 15.2 | 497 | 61 | 401 | 1.8 | 180.0 | −222 | 21.72 | 56.0 |
| 12006 | A. grandis | Eagle Creek—low USA/WA * | 760 | 120.6° | 47.7° | −4.2 | 7.6 | 17.8 | 831 | 56 | 531 | -0.8 | 185.0 | −352 | 19.05 | 46.0 |
| 12008 | A. grandis | Jack Creek USA/WA * | 825 | 120.8° | 47.3° | −3.7 | 6.6 | 15.9 | 877 | 56 | 487 | 1.1 | 185.0 | −308 | 17.30 | 43.0 |
| 12011 | A. grandis | Clear Lake USA/WA * | 945 | 121.3° | 46.6° | −3.8 | 5.4 | 14 | 948 | 69 | 394 | 3.0 | 172.0 | −215 | 17.89 | 48.0 |
| 12013 | A. grandis | Cooper Spur USA/OR | 1040 | 121.7° | 45.5° | −1.9 | 6.7 | 14.8 | 2097 | 115 | 232 | 2.2 | 126.0 | -53 | 18.91 | 36.0 |
| 12015 | A. grandis | Sisi Butte USA/OR * | 975 | 121.8° | 44.9° | −0.8 | 7.4 | 15.2 | 1595 | 98 | 318 | 1.8 | 143.0 | −139 | 18.19 | 36.0 |
| 12016 | A. grandis | Santiam Summit USA/OR | 1400 | 121.9° | 44.4° | −2.2 | 5.5 | 13.1 | 2033 | 127 | 266 | 3.9 | 114.0 | −87 | 16.22 | 34.0 |
| 12019 | A. grandis | Roaring River USA/OR * | 1310 | 122.0° | 43.5° | −0.7 | 6.8 | 14.3 | 1733 | 129 | 286 | 2.7 | 112.0 | −107 | 17.88 | 24.0 |
| 12020 | A. grandis | Crescent Creek USA/OR * | 1375 | 121.9° | 43.5° | −2.7 | 5.7 | 13.6 | 782 | 56 | 524 | 3.4 | 185.0 | −345 | 17.90 | 24.0 |
| 12025 | A. grandis | Buckskin Creek USA/ID * | 1220 | 116.2° | 48.0° | −5.4 | 4.8 | 14.4 | 1087 | 129 | 304 | 2.6 | 112.0 | −125 | 14.55 | 56.0 |
| 12026 | A. grandis | Plummer Hill USA/ID | 850 | 116.9° | 47.3° | −3.2 | 7.6 | 17.3 | 642 | 80 | 511 | -0.3 | 161.0 | −332 | 16.96 | 20.0 |
| 12031 | A. grandis | Bertha Hill USA/ID * | 1430 | 115.8° | 46.8° | −4.4 | 5.7 | 15.5 | 1341 | 120 | 307 | 1.5 | 121.0 | −128 | 18.17 | 63.0 |
| 12038 | A. grandis | Clearwater USA/ID * | 760 | 115.4° | 46.6° | −3.5 | 7.9 | 18.3 | 965 | 114 | 479 | −1.3 | 127.0 | −300 | 21.48 | 72.0 |
| 12037 | A. grandis | Stanley Creek USA/MT * | 800 | 115.9° | 48.3° | −5.4 | 6.1 | 16.1 | 698 | 97 | 494 | 0.9 | 144.0 | −315 | 18.58 | 53.0 |
| Data and current climate of the test site ** | 456 | 13.64° E | 49.79° | −1.4 | 8.1 | 17.0 | 595 | 241 | 179 | 19.26 | 50.1 | |||||
| Populations of the Provenance trial 219 Dražičky, CZ | Past Climate at Origin (1911–1940) | Climate Distance (D) | Data 2015 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Prov No. | Abies Species Name | Provenance Origin, Name | Alt. (m) | Long. W | Lat. N | January Mean Temp. (°C) | Annual Mean Temp. (°C) | Mean Temp. Warmest Quarter (°C) | Annual Mean Prec. (mm) | Mean Prec. Warmest Quarter (mm) | CMD (mm, ann.) | Temp. Change Warmest Quarter (°C) | Prec. Change Warmest Quarter (mm) | dCMD (Index Change, mm) | Mean Height (m) | Mean Tree Density (% from Planted) |
| CZ 0 | A. alba | Adršpach CZ | 620 | 16.1°E | 50.6° | −5.1 | 6.2 | 14.6 | 590 | 231 | 137 | 2.7 | −2.0 | 52.0 | 6.22 | 32.0 |
| 12001 | A. grandis | Buck Creek (Skamania) USA/WA | 400 | 121.4° | 48.3° | −0.6 | 8.5 | 16.5 | 2291 | 174 | 199 | 0.8 | 55.0 | −10.0 | 20.65 | 74.0 |
| 12002 | A. grandis | Tulalip (Ellensburg) USA/WA | 30 | 122.3° | 48.1° | 3.4 | 10.2 | 16.1 | 909 | 111 | 296 | 1.2 | 118.0 | −107.0 | 19.75 | 66.0 |
| 13004 * | A. procera | Mary’s Peak USA/OR | 1065 | 123.6° | 44.5° | 2.8 | 9.6 | 16.1 | 3060 | 89 | 269 | 1.2 | 140.0 | −80.0 | 12.41 | 44.0 |
| 13006 * | A. procera | Snow Peak USA/OR | 1060 | 122.6° | 44.6° | 1.5 | 7.9 | 14.3 | 2532 | 173 | 177 | 3.0 | 56.0 | 12.0 | 13.68 | 42.0 |
| 13011 * | A. procera | Larch Mtn. USA/OR | 975 | 122.1° | 45.5° | −0.3 | 7.2 | 14.2 | 2969 | 209 | 157 | 3.1 | 20.0 | 32.0 | 14.42 | 68.0 |
| 13014 * | A. procera | Red Mtn. USA/WA | 1220 | 121.8° | 45.9° | −2.4 | 5.4 | 13 | 2489 | 150 | 171 | 4.3 | 79.0 | 18.0 | 14.53 | 63.0 |
| 13018 | A. procera | McKinley Lake USA/WA | 900 | 122.1° | 46.6° | −0.4 | 7 | 14.2 | 2175 | 184 | 162 | 3.1 | 45.0 | 27.0 | 15.16 | 64.0 |
| 13021 * | A. procera | Stevens Pass USA/WA | 1000 | 121.1° | 47.7° | −3.8 | 5.1 | 13.2 | 2075 | 159 | 185 | 4.1 | 70.0 | 4.0 | 14.00 | 50.0 |
| Data and current climate of the test site ** | 488 | 14.59° E | 49.39° | −1.8 | 8.1 | 17.3 | 592 | 229 | 189 | 14.54 | 55.9 | |||||
| Location, Elevation (m), Geogr. Coordinates (Decimal Degrees) | Climate Period | Annual Mean Temperature (°C) | Anual Mean Precipitation (mm) | Annual CMD (mm) |
|---|---|---|---|---|
| Jíloviště CZ, Trial No.211 | 1911–1940 | 9 | 416 | 331 |
| 310 m | 1981–2010 | 9.2 | 477 | 296 |
| Lat. 49.944, Long. 14.356 | 2070–2100 | 13.5 | 522 | 389 |
| Sopron HU, comp.85 K. | 1911–1940 | 9.3 | 539 | 312 |
| 340 m | 1981–2010 | 9.5 | 623 | 276 |
| Lat. 47.667, Long. 16.563 | 2070–2100 | 14.1 | 661 | 393 |
| Zgornje Gruskovje SI, | 1911–1940 | 9.8 | 831 | 202 |
| 325 m | 1981–2010 | 10 | 965 | 172 |
| Lat. 46.272, Long. 15.860 | 2070–2100 | 14.6 | 978 | 271 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mátyás, C.; Beran, F.; Dostál, J.; Čáp, J.; Fulín, M.; Vejpustková, M.; Božič, G.; Balázs, P.; Frýdl, J. Surprising Drought Tolerance of Fir (Abies) Species between Past Climatic Adaptation and Future Projections Reveals New Chances for Adaptive Forest Management. Forests 2021, 12, 821. https://doi.org/10.3390/f12070821
Mátyás C, Beran F, Dostál J, Čáp J, Fulín M, Vejpustková M, Božič G, Balázs P, Frýdl J. Surprising Drought Tolerance of Fir (Abies) Species between Past Climatic Adaptation and Future Projections Reveals New Chances for Adaptive Forest Management. Forests. 2021; 12(7):821. https://doi.org/10.3390/f12070821
Chicago/Turabian StyleMátyás, Csaba, František Beran, Jaroslav Dostál, Jiří Čáp, Martin Fulín, Monika Vejpustková, Gregor Božič, Pál Balázs, and Josef Frýdl. 2021. "Surprising Drought Tolerance of Fir (Abies) Species between Past Climatic Adaptation and Future Projections Reveals New Chances for Adaptive Forest Management" Forests 12, no. 7: 821. https://doi.org/10.3390/f12070821