Microsites Influence the Light Response of Young Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco)


1
Slovenian Forestry Institute, Večna pot 2, 1000 Ljubljana, Slovenia
2
Department of Silviculture, Faculty of Forestry and Wood Technology, Mendel University Brno, Zemedelska 1665/1, 613 00 Brno, Czech Republic
Forests 2021, 12(6), 687; https://doi.org/10.3390/f12060687
Submission received: 3 May 2021
/
Revised: 25 May 2021
/
Accepted: 25 May 2021
/
Published: 27 May 2021
(This article belongs to the Section Forest Ecology and Management)
Abstract
:Two comparable sites with uneven-aged mixed forest stands with more than 20% Douglas-fir in the growing stock at an altitude of 650 m a.s.l. were selected. The physiological response of young trees to different light intensities was measured during the main growing season in three consecutive years, and four different light categories, which were determined from hemispherical photographs. The four light intensity categories were defined according to Indirect Site Factor (ISF%): in the open (A-ISF > 35%), at the outer forest edge (B-25% < ISF < 35%), at the inner forest edge (C-15% < ISF < 25%), and under complete canopy under mature forest stand (D-ISF < 15%). Climate data were obtained from the Royal Netherlands Meteorological Institute ‘Climate Explorer’ website. For the intensive micrometeorological observations, four monitoring sites were established along the elevation gradient at each site during summer and late fall to record relative humidity (RH%) and temperature (°C) with a 30 min recording interval sequence. Measured assimilation responses (A) and light use efficiency (Φ) at one site followed the expected pattern, while humidity combined with microsite conditions proved significant in explaining the specific response of young Douglas-fir to the different light intensity at the other site. For higher survival and optimal future development of Douglas-fir in the changing environment, microsites with higher capacity for storage moisture and favorable microclimate should generally be preferred to exposed and dry sites.
1. Introduction
The increasing frequency of extreme weather events and the associated larger amount of damaged forest areas have led to an impairment of the natural regeneration of autochthonous tree species. The selection of suitable tree species and planting concepts which could improve the resilience and resistance of future forest stands after such extreme events is questionable [1]. In discussions on the suitability of tree species, Douglas-fir (Pseudotsuga menziesii) is often suggested as a tree species that is more resilient to pests, drought, and weather extremes compared to the Norway spruce. Its role is assumed to be mainly as a substitute for declining spruce, which is threatened by climatic changes at lower altitudes [2], leading to a likely decrease in the supply of spruce wood in the near future [3].
The advantageous properties of Douglas-fir [4,5] are associated with a growth potential that is significantly higher than that of other resinous species [6], as well as its significant ecological plasticity. The species is known for its height (up to 75 m), high wood quality, and rapid growth [7,8]. It needs high humidity, and deep, fresh, and airy soils that can be acidified [9], but does not do well on limestone, accumulated, shallow, or poor soils with stagnant water [10,11,12]. It tolerates moderate shade, especially in juvenile stages, while its light requirement increases with age and retains its responsiveness longer than spruce; its plastic root system makes it better adapted to snowbreak and windthrow [13]. Height growth is slower in the first years and gradually increases to 1.2 m/year, which is maintained over several years. As a long-lived tree species, it can reach an enviable 1000 years [14] and maintain its regeneration under the mature canopy [15], but is sensitive to late frost events [16,17].
For Slovenian conditions, planting in mountain forests at altitudes between 500 and 1000 m above sea level has been recommended. The choice of provenance is important—for Slovenian conditions, the green Douglas-fir (Pseudotsuga menziesii subsp. Viridis) is the most suitable. It is also susceptible to fungal attack (Armilaria spp., Heterobasidium annosum, Phytopthora spp.…). The desirable mechanical properties of the wood, due to its excellent strength-to-weight ratio and higher density than other conifers, ensure its wide use [18]. The sustainability and resilience of the wood is much greater than that of spruce or fir, and does not require additional protection, therefore fetching much higher prices than other conifers [19]. The additional usefulness of needles, knots, resins, and bark is evident by the pleasant balsamic fragrance. Douglas-fir wood density, modulus of elasticity, and breaking and impact bending strength are higher than spruce or Scots pine [18].
Due to the great genetic variability of the Douglas-fir, it is suitable for introduction or further selection due to the diverse microsites in the local environment [20]. Previously, it was one of the most frequently planted tree species in Slovenia—until 1953, 17 objects with a total area of 16 ha were established; later, approximately 30 vital seed stands, well adapted to the specific Slovenian conditions, were registered. Due to the planned growth, Douglas-fir was planted in places with different locations and conditions (Josipdol—Ribnica na Pohorju, Rdeči breg—Podvelka, Počivalnik—Planina, Pečovnik—Celje, Rudnica—Podčetrtek, Jablje—Trzin, Brežice, Breginjski kot, Brkini,... etc.) [21].
The most important silvicultural tool to promote natural regeneration is a temporally and spatially appropriate stand structure and light environment to optimize the relationship between the future stand and site conditions. This is species and site specific, so it is important to know the response of different tree species to different light regimes, especially under changing environmental conditions. Light use efficiency (Φ) proved to be efficient in determining the competitive strength between silver fir and beech on different microsites in the high Dinaric karst [22], in comparing different silvicultural systems [23], as a response trait after large-scale disturbances [24], and in determining the optimal sites within a larger geographical region [25].
At two selected sites with abundant natural regeneration, where Douglas-fir has been successfully introduced in the past, we aimed to compare the species’ response in different light categories (1), and relate the response to influencing environmental parameters (2).
2. Material and Methods
2.1. Study Site and Data Collection
Surveys were conducted in the Celje (Celje local unit, sections 35343C, 35342B) and Postojna (Planina local unit, sections 01A30B, 01A30C) forest regional units of Slovenia Forest Service, both between 600 and 685 m a.s.l., with at least 20% Douglas-fir in the growing stock (Slovenia Forest Service). At each site, a contiguous area of established natural regeneration was selected under comparable light conditions with respect to the shading of the mature canopy stand. The predominant soil type in both cases was rendzic leptosol on carbonate bedrock. Site and plot characteristics are shown in Figure 1 and Table 1.
To obtain the best climate data for selected microsites, data on the mean monthly temperatures and total monthly precipitation were interpolated for the 0.5° grids that comprised each sampled stand and corresponded to the CRU TS 4.01 dataset [26]. Climate data were obtained from the Royal Netherlands Meteorological Institute ‘Climate Explorer’ website (http://climexp.knmi.nl, accessed on 21 March 2021).
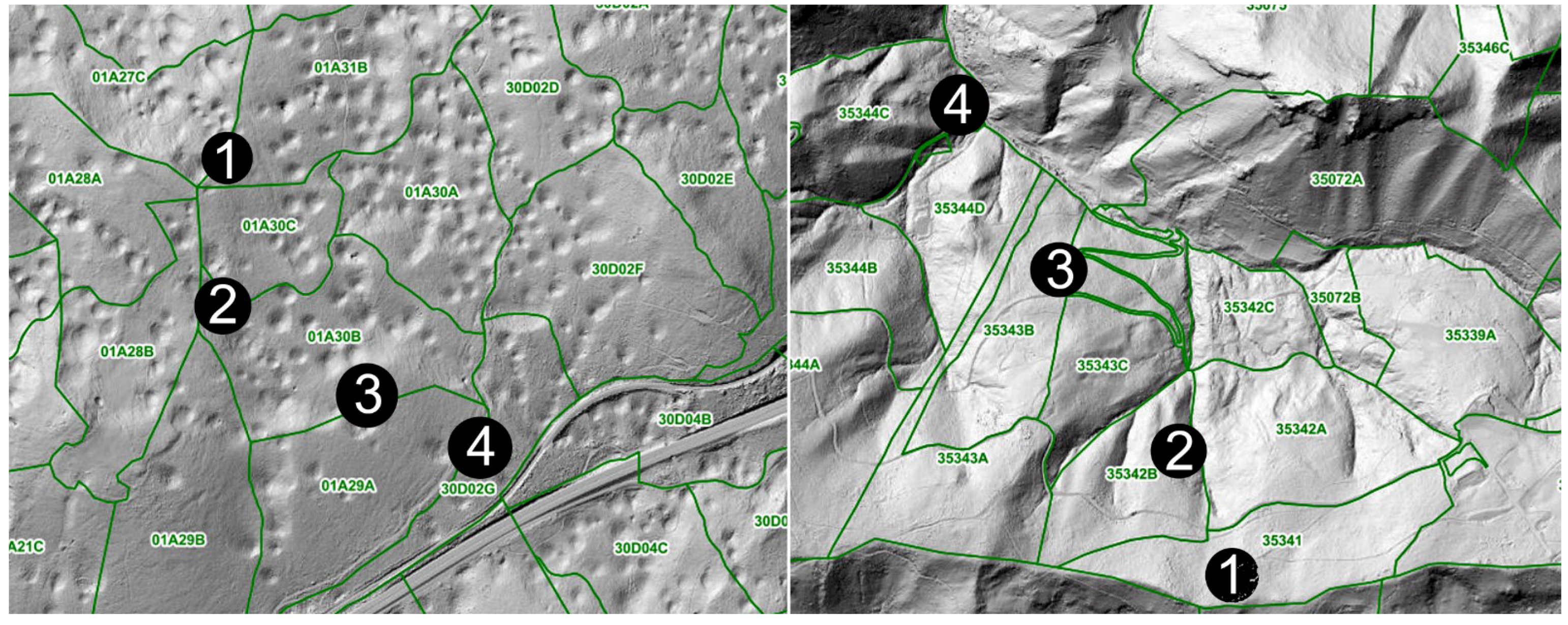
For the intensive micrometeorological observations, four measurement plots were established at each site (Figure 2) during summer and late fall to record relative humidity (RH%) and temperature (°C) with a 30 min recording interval sequence.
Categories of different light intensity were created based on the analysis of hemispherical photographs, taken with the Canon EOS Rebel T3i DSLR digital camera and a calibrated fisheye lens with the Regent WinScanopy accessory prior to physiological measurements during the main growing season. Light intensity parameters were processed using WinScanopy pro-d software [27]. Five hemispherical photographs were taken on each plot and light category. Based on the results, four light intensity categories were defined according to the Indirect Site Factor (ISF%) [22]: in the open (O-ISF > 35%), at the outer forest edge (Eo-25% < ISF < 35%), at the inner forest edge (Ei-15% < ISF < 25%), and under complete canopy cover beneath mature forest stand (S-ISF < 15%). Tree height ranged from 1.8 m under canopy to 3.2 m in the open light conditions.
Needles were collected from the upper canopy position of 8 trees per light category and site, and then stored in a cool, airtight environment. Nitrogen concentration (Ntot) [mg/g] was determined to compare macronutrient status (Leco CNS-2000 analyzer, St. Joseph, MI, USA) in all light categories. The same trees were used for assimilation response measurements.
Assimilation response (A) was measured during the main growing season in June and July for three consecutive years (2018–2020) from 7–11:30 a.m. during measurement campaigns. Trees measured were of comparable age (10–15 years) and size; in each light category at least 8 randomly selected trees were measured in the upper third of the crown each year using the LI-6400 portable infrared gas analyzer (IRGA) (Lincoln, NE, USA) with an autonomous light source under constant and computer-controlled environmental conditions. Net assimilation (A) rate in young trees to different light intensities and under comparable light conditions was measured at a constant temperature of the measurement block (20 °C), a CO2 concentration of 400 µmol/mol, a flux of 500 µmol/s, and different light intensities: 0, 50, 100, 250, 600, 1200 and 1800 µmol/m2s. All assimilation values were recorded after being held constant for 2 min or until the coefficient of variation (CV%) fell below 5%. Light use efficiency (Φ), defined as the amount of CO2 fixed per amount of light quanta absorbed (slope of the initial linear part of the light response curve) was determined for each light category (O, Eo, Ei and C) [25].
2.2. Data Analysis
Differences in light parameters (GF%, DSF%, ISF%), Ntot, A, and Φ in equal light categories between two selected sites were tested with factorial analyses of variance (ANOVA) after normality test and homogeneity of variances using the Statistica software system (2011). No transformation of the data was required to meet the criteria of normality in all tested parameters. Probability values of p < 0.05 (*), p < 0.01 (**) and p < 0.001 (***) were considered significant.
3. Results
3.1. Light Environment
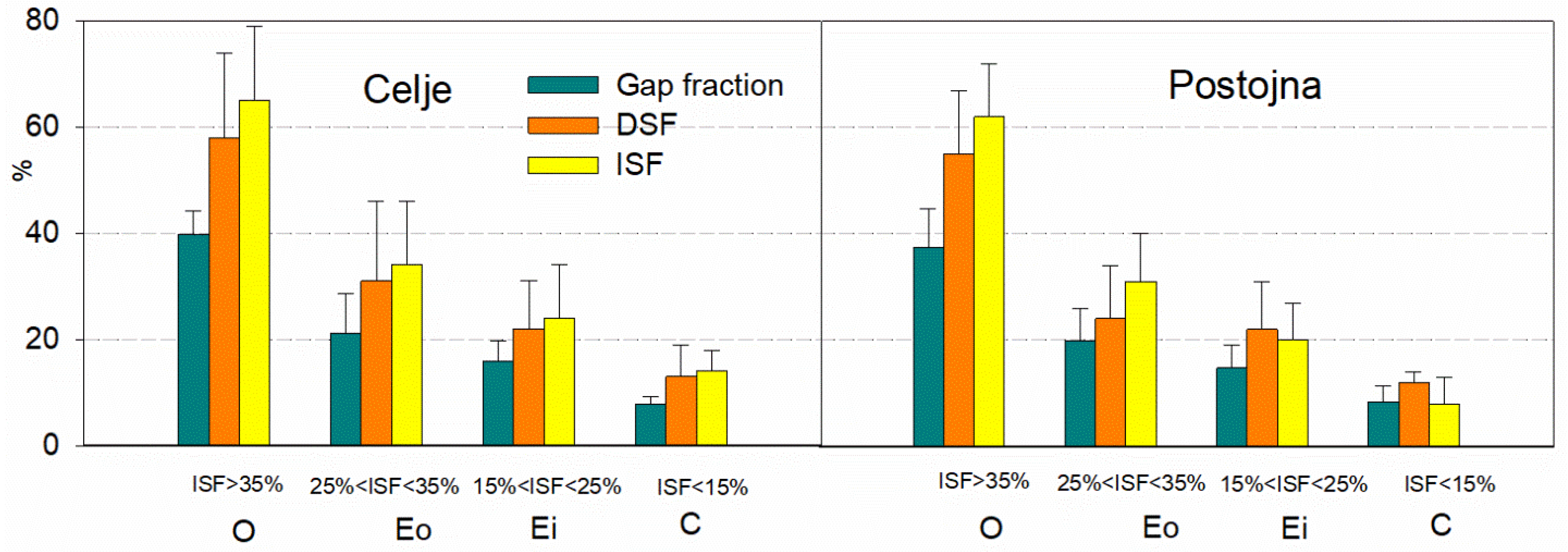
Hemispherical photo analysis confirmed comparable light parameters (GF-gap fraction; DSF-direct site factor; and ISF-indirect site factor) for the same light categories at both sites (df 1, 18). Light intensity decreases from open area conditions (O) to complete closure under mature stands (C) (Figure 3).
3.2. Nutrient Status
The foliar nitrogen content (Ntot) was highest in the open and lowest under canopy conditions at both sites, with no confirmed differences (AVAR, df 1, 18) between the same light categories at the different sites for all light parameters examined. The content was found within the optimum limits [28] (Table 2).
3.3. Climate and Micrometeorological Conditions
The average cumulative precipitation was higher in Postojna, while average air temperatures were higher in Celje for both long-term (1900–2019) and shorter, 10-year periods (Figure 3). Similar dynamics of changing parameters can be observed at both sites—an increase in average temperatures in the first four months (January–April) and midsummer (July–August), and increased precipitation redistribution, during the last three decades compared to long-term average conditions (Figure 4).
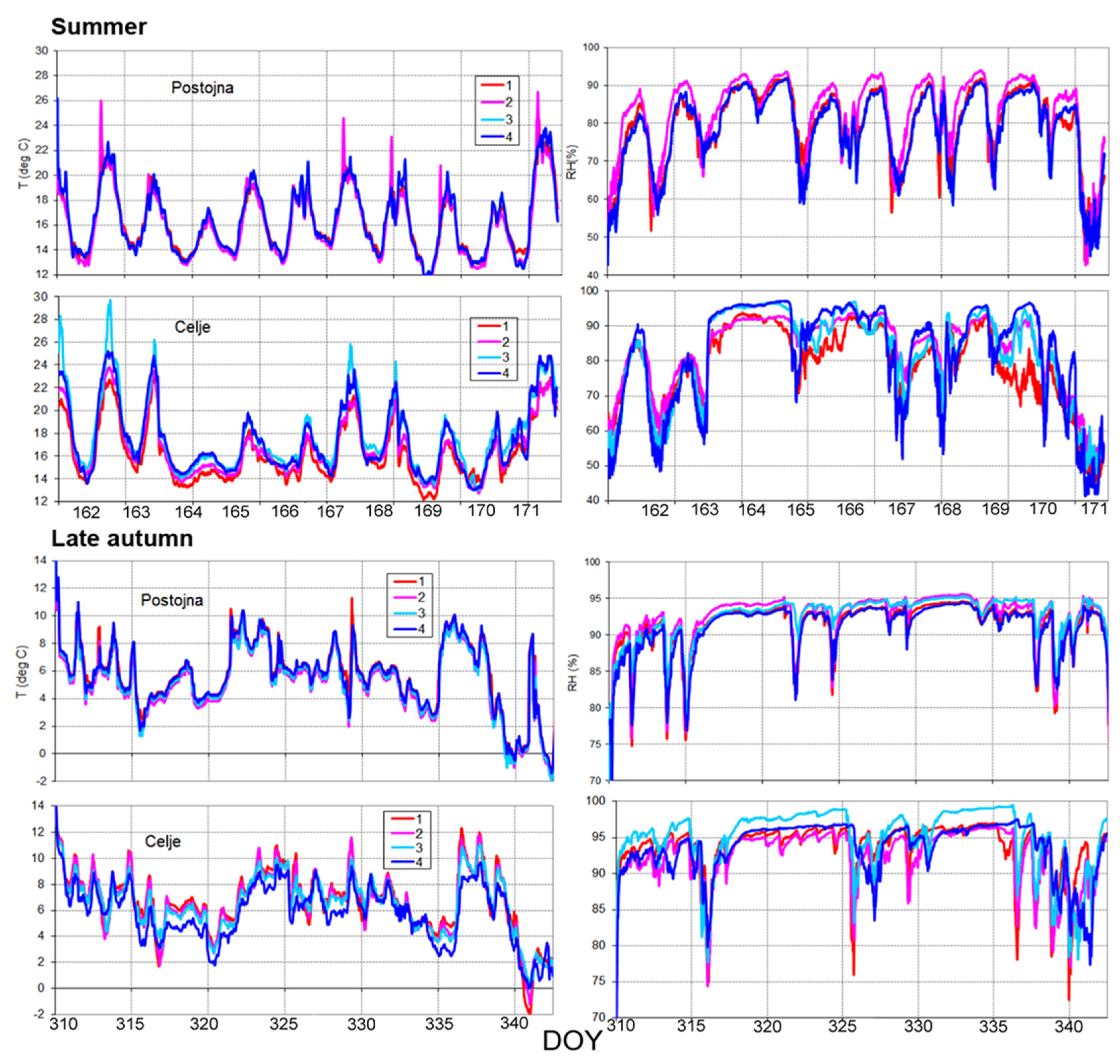
During the observation periods in summer (11.6.–22.6.2020) and late autumn (6.11.–5.12.2020), more homogeneous parameters were observed and measured at the Postojna site; higher values and greater dispersion of relative humidity (RH%) within the measured microsites were observed in Celje (Figure 5, Table 1).
Observed and measured conditions at the Postojna microsites followed the expected pattern from the highest RH (%) values on the top (1) to the lowest values on the bottom of the selected microsites (4). Humidity at the micro-sites in Celje was different—it was measured lowest at the highest point (1) and increased at lower elevations (4) (Figure 5). Not only the distribution, but the dispersion of both temperature and humidity between the four microsites was obviously larger in Celje.
In summer and late autumn, the average values of temperature and RH (%) were higher in Celje than in Postojna. The detailed values are presented in Table 3.
3.4. Assimilation
In Postojna, as expected, the assimilation rate (A) was highest in the open (O), followed by the outer forest edge (Eo), inner forest edge (Ei), and lowest under the complete canopy closure (C). In Celje, the response was different: the assimilation rate was highest in the of outer forest edge category (Eo), followed by the open category (O), the inner forest edge (Ei), and complete shelter (C). Absolute values in all comparable categories, except in the open without shelter (O), were highest in all cases in Celje (Figure 6). Responses between different years were similar, without significant differences.
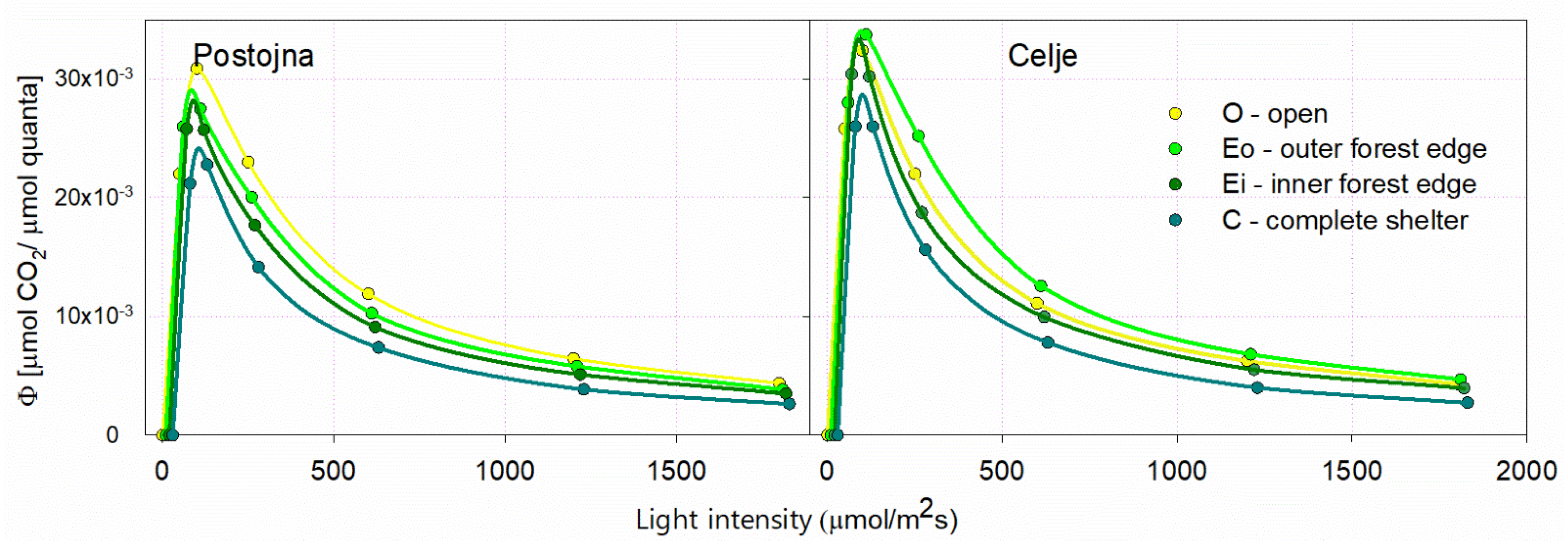
For light use efficiency (Φ), the relative distribution of light intensity categories was the same as that of the assimilation rate (A). In all comparable light categories, the light use efficiency was highest in Celje (Figure 7). Differences in A and Φ between years, sites and light categories are presented in Table 4.
4. Discussion
The impact of Douglas-fir on native systems is still not fully understood. As the oldest Douglas-fir stands currently growing in Europe have only reached approximately a fifth of the species’ maximum lifespan in its natural habitat, growth and interaction dynamics of mature, old-growth stands in Europe are still unknown [29].
Several studies have been conducted worldwide to evaluate the productivity of Douglas-fir sites as a function of environmental variables [6,30,31,32,33,34,35,36], and climatic conditions in particular. The high potential of Douglas-fir in mountainous regions has been recognized due to its growth rate, its ability to grow under a wide range of conditions, and its ability to regenerate naturally [37]. The site factors that most influence Douglas-fir growth are precipitation, soil depth, and elevation. The best production was found in regions with an average rainfall of more than 700 mm [38].
The average values of climatic parameters for the last three decades show a significant increase in temperature and a redistribution of precipitation, especially during the growing season (April–September). For both studied sites, there is a significant decrease in cumulative precipitation during 2000–2009, and an increased oscillation of precipitation compared to the long-term average distribution (1900–2019); the number of oscillations during the last three decades is increasing (Figure 3). In our study, absolute responses in all categories were highest in Celje, with slightly lower cumulative precipitation and higher average temperatures (according to KNMI Meteo database), compared to Postojna.
Foliar nitrogen in Douglas-fir is mainly influenced by soil water availability, and less so by other environmental factors [39]. In our case, nitrogen levels in needles (Ntot) confirmed no differences between the two sites and were found within the lower reported optimal range [39], indicating adequate water supply at both sites studied. Relatively modest values of Ntot at the rich sites studied could be related to the increase in temperature over the last three decades [28].
It appears that relative humidity affects young Douglas-fir performance much more than the absolute rainfall, at least at the sites studied. Together with somewhat lower Ntot values in all comparable light categories, one would expect higher absolute values from A and Φ at the Postojna site, but the results were just the opposite.
The measured assimilation responses in Postojna followed the expected pattern of light intensity categories from the most exposed in the open (O) to the lowest responses under shelter (C). The response in Celje was different—the assimilation response was highest in the outer forest edge category (Eo) and similar in the other categories. Despite the lower average temperature and higher precipitation in Postojna and optimal Ntot at both sites, the different response could be due to the slightly higher humidity in Celje in both midsummer and late autumn, and to the higher air temperatures in both periods (Table 2).
Douglas-fir is often considered to be a moderately shade-tolerant tree species, more light-requiring than the Norway spruce and silver fir [40]. Our study confirms such light-adapted character of young trees, as the maximum Φ was measured at the outer forest edge (Eo), in contrast to fir, where the maximum Φ values from other studies were measured under shelter, and decreased with increasing light [22].
Several authors report that the Douglas-fir’s resistance to transient drought is physiologically attributed to a more effective control mechanism of the stomata than in other conifers [41,42]. In a nearly 600 km area along its natural distribution adjacent to Pacific Ocean, the coastal fog belt mitigates the repetitive pattern of several-month summer droughts, indicating a climate-induced drought adaptation of the species [43,44]. The configuration of the Celje site in the highlands, with a chain of ridges at elevations between 700 and 800 m, and a catchment with numerous tributaries [45], may contribute significantly to the site-specific response, causing pronounced retention of humidity/moisture and fog up to the level of the inversion layer, where naturally regenerated Douglas-fir is found. In contrast, the pronounced karst terrain at the Postojna site, which is located on the southern edge of Planinsko polje, and has sufficient moisture and water, leads to a different response in young trees with a more open and wind-exposed terrain than at the Celje site.
Soil analysis in the study conducted on acidic sites with natural hardwood, spruce monocultures and Douglas-fir stands showed favorable effects of this tree species on soil chemistry, organic matter, and nutrient dynamics. Compared to native conifer species, Douglas-fir had a lower acidifying effect on the upper soil layers, and contributed to better humus formation by recycling nutrients more effectively and producing litter that could be easily decomposed [46]. A significant difference was found in the allometric ratios of spruce from the nutrient-poor and nutrient-rich sites. In contrast to spruce, site fertility did not confirm a significant effect on allometric ratios in Douglas-fir, suggesting that their allocation patterns are less sensitive to site nutrients [47]. Douglas-fir stands influence their habitats, as shown by changes in species composition in the ground vegetation, and the abundance and dominance of certain species on forest sites in Central Europe [48]. Despite the favorable results, we do not have similar long-term comparisons in Slovenia that would confirm or refute the stated benefits. Another challenge is the large proportion of Natura 2000 sites, which cover more than 45% of forests (Slovenia Forest Service 2019). Management in these areas is subject to stricter criteria where the introduction of non-native tree species, including Douglas-fir, is prohibited.
According to Sauter [19], Douglas-fir can achieve an average annual growth of 19.3 m3/ha and heights of up to 75 m [49], which is more than any of the dominant tree species in Europe. Air temperatures, last year’s precipitation deficit, and their interactions are the main factors affecting growth in all habitats. Winter temperatures are more important at higher and colder altitudes, just as summer temperatures are in warm and dry habitats. Due to climate change, growth has been increasing at higher elevations, and decreasing at temperate ones, since 1900 [50]. Radial growth is influenced by the local microclimate—it is positively correlated with drought index and summer and autumn precipitation of the previous growing season, confirming the influence of soil water supply. Annual precipitation has a pronounced seasonal effect, and drought stress due to lower soil water supply is the predominant climatic factor affecting radial growth [51]. Positive correlations of growth with monthly temperatures reflect the positive effect of warmer periods on photosynthesis and radial growth during periods of sufficient soil water supply [51], which is also confirmed by our results, and the fact that at the same site in Celje, groups of mature Douglas-fir were measured in the category of the tallest trees in Slovenia (67 m) [52]. Unfortunately, comparison with the Postojna site was not possible, as there were no trees at this site of the same age as those in Celje.
Humidity associated with microsite conditions proved to be significant in explaining the specific response of young Douglas-fir with the highest values under lightly shaded conditions (Celje), compared to other sites where the response followed the expected light intensity categories. In the future, a more substantial impact of the specific microsite effect can be expected if the increase in temperatures during the growing season follows the reported pace of the last decades. The possibility of replacing spruce at lower altitudes below 700 m a.s.l. and the admixture of Douglas-fir with other tree species may represent a realistic and cost-effective alternative, which would reduce the risk for better drought tolerance and better mechanical properties on suitable sites [53,54]. However, the recommendation as an alternative tree species for declining spruce at lower altitudes should be regarded with caution, as Douglas-fir was also revealed to have increased sensitivity to increased temperatures and drought stress in Central Europe [55]. For a higher survival rate and optimal future development of Douglas-fir in the changing environment, microsites with higher capacity for retention moisture and favorable microclimates should generally be preferred over exposed and dry sites.
Funding
The author gratefully acknowledges the financial support from the Slovenian Research Agency (Research Core Funding No. P4-0107 Program Research Group ‘‘Forest Biology, Ecology and Technology” at Slovenian Forestry Institute, and the Ministry of Agriculture, Forestry and Food (CRP projects V4-1820 and V4-1818).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Publicly available datasets were analyzed in this study. The data presented in this study are available on request from the corresponding author.
Acknowledgments
Mag. R. Hostnik and the team from the Slovenia Forest Service, Celje unit helped me with site selection.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Brang, P.; Spathelf, P.; Larsen, J.B.; Bauhus, J.; Bončìna, A.; Chauvin, C.; Drössler, L.; García-Güemes, C.; Heiri, C.; Kerr, G. Suitability of close-to-nature silviculture for adapting temperate European forests to climate change. For. Int. J. For. Res. 2014, 87, 492–503. [Google Scholar] [CrossRef] [Green Version]
- Podrázský, V.; Remeš, J.; Sloup, R.; Pulkrab, K.; Novotná, S. Douglas-fir-Partial substitution for declining conifer timber supply-review of Czech data. Wood Res. 2016, 61, 525–530. [Google Scholar]
- Synek, M.; Vašíček, J.; Zeman, M. Outlook of logging perspectives in the Czech Republic for the period 2013–2032. J. For. Sci. 2014, 60, 372–381. [Google Scholar] [CrossRef] [Green Version]
- Henin, J.-M.; Pollet, C.; Jourez, B.; Hébert, J. Impact of Tree Growth Rate on the Mechanical Properties of Douglas Fir Lumber in Belgium. Forests 2018, 9, 342. [Google Scholar] [CrossRef] [Green Version]
- Pollet, C.; Henin, J.-M.; Hébert, J.; Jourez, B. Effect of growth rate on the physical and mechanical properties of Douglas-fir in western Europe. Can. J. For. Res. 2017, 47, 1056–1065. [Google Scholar] [CrossRef]
- Hein, S.; Weiskittel, A.R.; Kohnle, U. Effect of wide spacing on tree growth, branch and sapwood properties of young Douglas-fir [Pseudotsuga menziesii (Mirb.) Franco] in south-western Germany. Eur. J. For. Res. 2008, 127, 481–493. [Google Scholar] [CrossRef]
- Bergel, D. Douglasien-Ertragstafel für Nordwestdeutschland. In Ertragstafeln Wichtiger Baumarten; Schober, R., Ed.; J. D. Sauerländer’s Verlag, 3. neu bearbeitete und erweiterte Auflage: Frankfurt am Main, Germany, 1985. [Google Scholar]
- Begin, J. Productivite du Douglas vert (Pseudotsuga menziesii (Mirb) Franco var menziesii) en Relation avec des Characteristiques Stationnelles. Ph.D. Thesis, ETH Zürich, Zürich, Switzerland, 1991. [Google Scholar]
- Jasser, C. Douglasie in Oberösterreich—Möglichkeiten und Grenzen; Bundesforschungs-und Ausbildungszentrum für Wald, Naturgefahren und Landschaft: Wien, Austria, 2008; pp. 19–20. [Google Scholar]
- Riehl, G. Zum Waldbau der Douglasie in Nordwestdeutschland—Erfahrungen eines Praktikers. Forst Holz 2000, 55, 716–718. [Google Scholar]
- Englisch, M. Die Douglasie—Für und Wider aus Standortkundlicher Sicht; Bundesforschungs-und Ausbildungszentrum für Wald, Naturgefahren und Landschaft: Wien, Austria, 2008; pp. 6–8. [Google Scholar]
- Meyer, M.; Ulrich, B. Auswirkungen einer Kalkung auf Böden mit Mangantoxizität bei Douglasienbeständen auf Buntsandstein in der Nordeifel. Forst Holz 1900, 45, 493–498. [Google Scholar]
- Albert, M.; Hansen, J.; Nagel, J.; Schmidt, M.; Spellmann, H. Assessing risks and uncertainties in forest dynamics under different management scenarios and climate change. For. Ecosyst. 2015, 2, 14. [Google Scholar] [CrossRef] [Green Version]
- Kuiper, L.C. The Structure of Natural Douglas-Fir Forests in Western Washington and Western Oregon; Agricultural University Wageningen Papers; Agricultural University Wageningen: Wageningen, The Netherlands, 1988; Volume 88, p. 47. [Google Scholar]
- Knoerzer, D.; Reif, A. Die Naturverjüngung der Douglasie im Bereich des Stadtwaldes von Freiburg. Afz-Der Wald 1996, 51, 1117–1120. [Google Scholar]
- Lévesque, M.; Saurer, M.; Siegwolf, R.; Eilmann, B.; Brang, P.; Bugmann, H.; Rigling, A. Drought response of five conifer species under contrasting water availability suggests high vulnerability of Norway spruce and European larch. Glob. Chang. Biol. 2013, 19, 3184–3199. [Google Scholar] [CrossRef]
- Lévesque, M.; Rigling, A.; Bugmann, H.; Weber, P.; Brang, P. Growth response of five co-occurring conifers to drought across a wide climatic gradient in Central Europe. Agric. For. Meteorol. 2014, 197, 1–12. [Google Scholar] [CrossRef]
- Zeidler, A.; Borůvka, V.; Schönfelder, O. Comparison of Wood Quality of Douglas Fir and Spruce from Afforested Agricultural Land and Permanent Forest Land in the Czech Republic. Forests 2018, 9, 13. [Google Scholar] [CrossRef] [Green Version]
- Sauter, U. Characteristics and potential of Douglas fir (Pseudotsuga Menziesii (Mirb) Franco); Forest Research Institute, Department of Forest Utilisation: Baden-Wurtttenberg, Germany, 2008; p. 38. [Google Scholar]
- Breznikar, A. Mednarodno provenienčno preučevanje duglazije (Pseudotsuga menziesii (Mirb) Franco) v Sloveniji- International Provenance Research on Douglas fir (Pseudotsuga Menziesii (Mirb.) Franco) in Slovenia; University of Ljubljana: Ljubljana, Slovenia, 1991. [Google Scholar]
- Čokl, M. Rast Tujih Iglavcev v Sloveniji; IGLG: Ljubljana, Slovenia, 1965; p. 234. [Google Scholar]
- Čater, M.; Diaci, J.; Roženbergar, D. Gap size and position influence variable response of Fagus sylvatica L. and Abies alba Mill. For. Ecol. Manag. 2014, 325, 128–135. [Google Scholar] [CrossRef]
- Čater, M.; Levanič, T. Response of Fagus sylvatica L. and Abies alba Mill. in different silvicultural systems of the high Dinaric karst. For. Ecol. Manag. 2013, 289, 278–288. [Google Scholar] [CrossRef]
- Čater, M.; Diaci, J. Divergent response of European beech, silver fir and Norway spruce advance regeneration to increased light levels following natural disturbance. For. Ecol. Manag. 2017, 399, 206–212. [Google Scholar] [CrossRef]
- Čater, M.; Levanič, T. Beech and silver fir’s response along the Balkan’s latitudinal gradient. Sci. Rep. 2019, 9, 16269. [Google Scholar] [CrossRef] [Green Version]
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations–the CRU TS3. 10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef] [Green Version]
- Čater, M.; Schmid, I.; Kazda, M. Instantaneous and potential radiation effect on underplanted European beech below Norway spruce canopy. Eur. J. For. Res. 2012, 132. [Google Scholar] [CrossRef]
- Lewis, J.D.; Lucash, M.M.; Olszyk, D.M.; Tingey, D.T. Relationships between needle nitrogen concentration and photosynthetic responses of Douglas-fir seedlings to elevated CO2 and temperature. New Phytol. 2004, 162, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Douglas-fir—An Option for Europe. EFI What Science Can Tell Us. In COST Action FP1403 NNEXT, Supported by COST (European Cooperation in Science and Technology); European Cooperation in Science and Technology: Brussels, Belgium, 2019; p. 124. Available online: https://efi.int/sites/default/files/files/publication-bank/2019/efi_wsctu9_2019.pdf (accessed on 21 March 2021).
- Tyler, A.L.; MacMillan, D.C.; Dutch, J. Models to predict the General Yield Class of Douglas fir, Japanese larch and Scots pine on better quality land in Scotland. For. Int. J. For. Res. 1996, 69, 13–24. [Google Scholar] [CrossRef]
- Curt, T.; Bouchaud, M.; Agrech, G. Predicting site index of Douglas-Fir plantations from ecological variables in the Massif Central area of France. For. Ecol. Manag. 2001, 149, 61–74. [Google Scholar] [CrossRef]
- Corona, P.; Scotti, R.; Tarchiani, N. Relationship between environmental factors and site index in Douglas-fir plantations in central Italy. For. Ecol. Manag. 1998, 110, 195–207. [Google Scholar] [CrossRef]
- Klinka, K.; Carter, R.E. Relationships between Site Index and Synoptic Environmental Factors in Immature Coastal Douglas-Fir Stands. For. Sci. 1990, 36, 815–830. [Google Scholar] [CrossRef]
- Monserud, R.A.; Moody, U.; Breuer, D.W. A soil-site study for inland Douglas-fir. Can. J. For. Res. 1990, 20, 686–695. [Google Scholar] [CrossRef]
- Dunbar, A.; Dhubhain, A.N.; Bulfin, M. The productivity of Douglas fir in Ireland. For. Int. J. For. Res. 2002, 75, 537–545. [Google Scholar] [CrossRef]
- Kubeček, J.; Štefančík, I.; Podrázský, V.; Longauer, R. Výsledky výzkumu douglasky tisolisté (Pseudotsuga menziesii/Mirb./Franco) v České republice a na Slovensku—Přehled. [Results of the research of Douglas-fir in the Czech Republic and Slovakia-review]. For. J. 2014, 60, 120–129. [Google Scholar]
- Fontes, L.; Tomé, M.; Thompson, F.; Yeomans, A.; Luis, J.S.; Savill, P. Modelling the Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) site index from site factors in Portugal. For. Int. J. For. Res. 2003, 76, 491–507. [Google Scholar] [CrossRef] [Green Version]
- Fontes, L.; Tomé, M.; Coelho, M.B.; Wright, H.; Luis, J.S.; Savill, P. Modelling dominant height growth of Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) in Portugal. For. Int. J. For. Res. 2003, 76, 509–523. [Google Scholar] [CrossRef] [Green Version]
- Du, B.; Kreuzwieser, J.; Dannenmann, M.; Junker, L.V.; Kleiber, A.; Hess, M.; Jansen, K.; Eiblmeier, M.; Gessler, A.; Kohnle, U.; et al. Foliar nitrogen metabolism of adult Douglas-fir trees is affected by soil water availability and varies little among provenances. PLoS ONE 2018, 13, e0194684. [Google Scholar] [CrossRef]
- Savill, P. The Silviculture of Trees Used in British Forestry, 2nd ed.; CABI Publishing: Wallingford, Oxfordshire, UK, 2013. [Google Scholar]
- Lassoie, J.P.; Salo, D.J. Physiological response of large Douglas-fir to natural and induced soil water deficits. Can. J. For. Res. 1981, 11, 139–144. [Google Scholar] [CrossRef]
- Grieu, P.; Guehl, J.M.; Aussenac, G. The effects of soil and atmospheric drought on photosynthesis and stomatal control of gas exchange in three coniferous species. Physiol. Plant. 1988, 73, 97–104. [Google Scholar] [CrossRef]
- Jenssen, M. Die Grüne Douglasie im klimaplastischen Wald des Tieflandes—Ökoklimatisch-waldgeografische Grundlagen. Eberswalder Forstl. Schr. 2009, 43, 15–22. [Google Scholar]
- Bansal, S.; Harrington, C.A.; Gould, P.J.; St Clair, J.B. Climate-related genetic variation in drought-resistance of Douglas-fir (Pseudotsuga menziesii). Glob. Chang. Biol 2015, 21, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Košutnik, D. Celjski Mestni Gozd v Pečovniku; Hostnik, R., Ed.; Zavod za Gozdove Slovenije: Ljubljana, Slovenije, 2013; p. 193. [Google Scholar]
- Kupka, I.; Podrázský, V.; Kubeček, J. Soil-forming effect of Douglas fir at lower altitudes—A case study. J. For. Sci. 2013, 59, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Urban, J.; Holušová, K.; Menšík, L.; Čermák, J.; Kantor, P. Tree allometry of Douglas fir and Norway spruce on a nutrient-poor and a nutrient-rich site. Trees 2013, 27, 97–110. [Google Scholar] [CrossRef]
- Podrázský, V.; Martiník, A.; Matejka, K.J.; Viewegh, J. Effects of Douglas-fir (Pseudotsuga menziesii [Mirb.] Franco) on understorey layer species diversity in managed forests. J. For. Sci. 2014, 60, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Remeš, J.; Zeidler, A. Production potential and wood quality of Douglas fir from selected sites in the Czech Republic. Wood Res. 2014, 59, 509–520. [Google Scholar]
- Lee, E.H.; Beedlow, P.A.; Waschmann, R.S.; Tingey, D.T.; Wickham, C.; Cline, S.; Bollman, M.; Carlile, C. Douglas-fir displays a range of growth responses to temperature, water, and Swiss needle cast in western Oregon, USA. Agric. For. Meteorol. 2016, 221, 176–188. [Google Scholar] [CrossRef]
- Little, R.L.; Peterson, D.L.; Silsbee, D.G.; Shainsky, L.J.; Bednar, L.J. Radial growth patterns and the effects of climate on second-growth Douglas-fir (Pseudotsuga menziesii) in the Siskiyou Mountains, Oregon. Can. J. For. Res. 1995, 25, 724–735. [Google Scholar] [CrossRef]
- Hostnik, R. Velikanke/The Giantess. In Najvišje Izmerjeno Drevo v Sloveniji/The Tallest Measured Tree in Slovenia; Zavod za gozdove Slovenije, Društvo za Trajnostni Razvoj Urbane Narave: Celje, Slovenia; Available online: https://www.youtube.com/watch?v=Cct0Nb5Yj_o (accessed on 21 March 2021).
- Podrázský, V.; Kupka, I.; Prknová, H. Substitution of Norway spruce for Douglas-fir: Changes of soil microbial activities as climate change induced shift in species composition—A case study. Cent. Eur. For. J. 2020, 66, 71–77. [Google Scholar] [CrossRef]
- Remeš, J.; Pulkrab, K.; Bílek, L.; Podrázský, V. Economic and Production Effect of Tree Species Change as a Result of Adaptation to Climate Change. Forests 2020, 11, 431. [Google Scholar] [CrossRef] [Green Version]
- Vejpustková, M.; Čihák, T. Climate Response of Douglas Fir Reveals Recently Increased Sensitivity to Drought Stress in Central Europe. Forests 2019, 10, 97. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Research site location.

Figure 2.
Location of micrometeorological stations, arranged from the locally highest (1) to the lowest (4) elevation at the Postojna (left) and Celje (right) research sites (M = 1:1000; source: Slovenia Forest Service).
Figure 2.
Location of micrometeorological stations, arranged from the locally highest (1) to the lowest (4) elevation at the Postojna (left) and Celje (right) research sites (M = 1:1000; source: Slovenia Forest Service).

Figure 3.
Light categories and different parameters from the hemispherical photo analysis at both sites. O-open; Eo- outer forest edge; Ei-inner forest edge and C-complete canopy cover. The bars are standard errors.
Figure 3.
Light categories and different parameters from the hemispherical photo analysis at both sites. O-open; Eo- outer forest edge; Ei-inner forest edge and C-complete canopy cover. The bars are standard errors.

Figure 4.
Average monthly temperature and precipitation during the long-term period (1900–2019) and the last three decades (1990–1999/2000–2009/2010–2019). Total annual precipitation and mean annual temperatures are shown near the corresponding period. Source: KNMI.
Figure 4.
Average monthly temperature and precipitation during the long-term period (1900–2019) and the last three decades (1990–1999/2000–2009/2010–2019). Total annual precipitation and mean annual temperatures are shown near the corresponding period. Source: KNMI.

Figure 5.
Dynamics of temperature (°C) and humidity (RH%) in summer (upper part) and late autumn (lower part) periods from the highest (1) to the lowest micro site (4).
Figure 5.
Dynamics of temperature (°C) and humidity (RH%) in summer (upper part) and late autumn (lower part) periods from the highest (1) to the lowest micro site (4).

Figure 6.
The assimilation rate (A) as a function of light intensity at the Celje (left) and Postojna (right) sites. Values shown are shifted to avoid overlap and refer to the same light intensity. The bars are standard errors.
Figure 6.
The assimilation rate (A) as a function of light intensity at the Celje (left) and Postojna (right) sites. Values shown are shifted to avoid overlap and refer to the same light intensity. The bars are standard errors.

Figure 7.
Light use efficiency (Φ) for light categories in Postojna and Celje.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Postojna and Celje site characteristics.
| Location | Latitude (N) | Longitude (E) | Elevation (m a.s.l.) | Growing Stock (m3) |
|---|---|---|---|---|
| Postojna | 45°48′29.6′′ | 14°15′33.0′′ | 657 | 515 |
| Celje | 46°11′32.2′′ | 15°15′51.8′′ | 651 | 540 |
Table 2.
Foliar nitrogen (Ntot; mg/g) for all light categories (O-open; Eo- outer forest edge; Ei-inner forest edge and C-complete canopy cover) and both sites (N = 8). Mean values with standard error of the mean are given.
Table 2.
Foliar nitrogen (Ntot; mg/g) for all light categories (O-open; Eo- outer forest edge; Ei-inner forest edge and C-complete canopy cover) and both sites (N = 8). Mean values with standard error of the mean are given.
| Site/Light Category | O | Eo | Ei | C |
|---|---|---|---|---|
| Postojna | 13.2 ± 3.1 | 13.1 ± 2.4 | 13.1 ± 2.9 | 12.8 ± 4.1 |
| Celje | 12.9 ± 4.2 | 12.9 ± 3.6 | 12.8 ± 3.9 | 12.8 ± 3.7 |
Table 3.
Average temperature and humidity values for summer and late autumn microsites in Postojna and Celje. Means and standard errors are shown.
Table 3.
Average temperature and humidity values for summer and late autumn microsites in Postojna and Celje. Means and standard errors are shown.
| Postojna | Celje | ||||
|---|---|---|---|---|---|
| Microsite | T (°C) | RH (%) | T (°C) | RH (%) | |
| DOY: 310-340 | 1 | 5.4 ± 2.7 | 90.7 ± 4.6 | 5.5 ± 2.7 | 96.7 ± 3.7 |
| 2 | 4.9 ± 2.6 | 91.7 ± 4.5 | 6.3 ± 2.9 | 99.5 ± 3.7 | |
| 3 | 5.0 ± 2.4 | 91.0 ± 3.8 | 6.2 ± 2.9 | 95.0 ± 4.3 | |
| 4 | 5.3 ± 2.3 | 90.5 ± 4.2 | 6.2 ± 2.7 | 94.6 ± 3.9 | |
| mean | 5.2 ± 2.6 | 91.0 ± 4.3 | 6.0 ± 2.8 | 96.5 ± 3.9 | |
| DOY: 162-171 | 1 | 16.2 ± 2.4 | 78.7 ± 9.8 | 15.2 ± 2.6 | 79.0 ± 11.0 |
| 2 | 16.0 ± 2.8 | 77.6 ± 10.0 | 16.8 ± 2.6 | 83.3 ± 10.2 | |
| 3 | 16.3 ± 2.7 | 78.7 ± 10.2 | 17.8 ± 3.2 | 82.1 ± 12.7 | |
| 4 | 16.4 ± 2.6 | 78.0 ± 10.1 | 17.4 ± 3.0 | 82.1 ± 14.9 | |
| mean | 16.2 ± 2.6 | 78.2 ± 10.0 | 17.0 ± 2.8 | 81.6 ± 12.2 | |
Table 4.
Differences in A and Φ between years, sites and light categories. Probability values of p < 0.001 (***) were considered significant; NS—non-significant.
Table 4.
Differences in A and Φ between years, sites and light categories. Probability values of p < 0.001 (***) were considered significant; NS—non-significant.
| A | Φ | ||||
|---|---|---|---|---|---|
| df | F | p | F | p | |
| Year | 2 | 0.2 | 0.84 NS | 0.9 | 0.41 NS |
| Site | 1 | 753.5 | 0.000 *** | 164.4 | 0.000 *** |
| Light category | 3 | 3781.4 | 0.000 *** | 1371.1 | 0.000 *** |
| Year × Site | 2 | 0.0 | 0.96 NS | 0.9 | 0.42 NS |
| Year × Light category | 6 | 0.1 | 0.99 NS | 0.3 | 0.94 NS |
| Site × Light category | 3 | 306.0 | 0.000 *** | 153.1 | 0.000 *** |
| Year × Site × Light category | 6 | 0.2 | 0.84 NS | 0.2 | 0.97 NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Čater, M. Microsites Influence the Light Response of Young Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco). Forests 2021, 12, 687. https://doi.org/10.3390/f12060687
AMA Style
Čater M. Microsites Influence the Light Response of Young Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco). Forests. 2021; 12(6):687. https://doi.org/10.3390/f12060687
Chicago/Turabian StyleČater, Matjaž. 2021. "Microsites Influence the Light Response of Young Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco)" Forests 12, no. 6: 687. https://doi.org/10.3390/f12060687
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.