Versatility of Synthetic tRNAs in Genetic Code Expansion


1
Department of Molecular Biophysics and Biochemistry, Yale University, New Haven, CT 06520, USA
2
Department of Chemistry, Yale University, New Haven, CT 06520, USA
*
Author to whom correspondence should be addressed.
Genes 2018, 9(11), 537; https://doi.org/10.3390/genes9110537
Submission received: 16 October 2018
/
Revised: 31 October 2018
/
Accepted: 5 November 2018
/
Published: 7 November 2018
(This article belongs to the Special Issue Synthetic DNA and RNA Programming)
Abstract
:Transfer RNA (tRNA) is a dynamic molecule used by all forms of life as a key component of the translation apparatus. Each tRNA is highly processed, structured, and modified, to accurately deliver amino acids to the ribosome for protein synthesis. The tRNA molecule is a critical component in synthetic biology methods for the synthesis of proteins designed to contain non-canonical amino acids (ncAAs). The multiple interactions and maturation requirements of a tRNA pose engineering challenges, but also offer tunable features. Major advances in the field of genetic code expansion have repeatedly demonstrated the central importance of suppressor tRNAs for efficient incorporation of ncAAs. Here we review the current status of two fundamentally different translation systems (TSs), selenocysteine (Sec)- and pyrrolysine (Pyl)-TSs. Idiosyncratic requirements of each of these TSs mandate how their tRNAs are adapted and dictate the techniques used to select or identify the best synthetic variants.

1. Introduction
Genetic code expansion (GCE) involves the engineering of protein synthesis machinery to site-specifically incorporate non-canonical amino acids (ncAAs) into a desired protein [1,2]. This is routinely done by assigning the ncAA to recoded stop or sense codons and delivering the ncAA to the ribosome via a suppressor transfer RNA (tRNA). The successful charging of an ncAA to the suppressor tRNA and incorporation at a defined codon requires an aminoacyl-tRNA synthetase (aaRS)•tRNA pair to function orthogonally (restricting interactions with host tRNAs, aaRSs, or canonical amino acids; Figure 1). Non-canonical amino acids endow proteins with unique chemical and physical properties that make them useful for a wide range of applications. They serve as affinity tags, imaging probes, environmental sensors, post-translational modifications, are used for protein crosslinking, conjugation, and altering pKa or redox potential [3].
The most versatile aaRS for incorporating ncAAs is pyrrolysyl-tRNA synthetase (PylRS). Naturally, PylRS attaches pyrrolysine (Pyl), the 22nd genetically encoded amino acid, to its cognate tRNAPyl, a natural UAG suppressor. In archaea, PylRS is a single polypeptide chain; however, bacteria harbor a split protein where the C-terminal catalytic domain is only active in the presence of the N-terminal domain [4,5]. PylRS and its variants are polyspecific; to date they have facilitated the incorporation of over 100 ncAAs into proteins [6]. Moreover, PylRS•tRNAPyl pairs are used to engineer proteins with unique properties and functions in bacteria, viruses, insects, yeast, and animals [7,8,9,10,11].
Another valuable building block for protein engineering is the 21st amino acid, selenocysteine (Sec). Sec is a naturally occurring amino acid that resembles cysteine but has a selenol group instead of the thiol. Sec is found in the active site of redox enzymes of species that span all three domains of life, providing enhanced nucleophilic and reducing properties [12]. The site-specific incorporation of Sec can enhance enzyme activity when replacing cysteine (Cys), increase protein stability via diselenide bonds, and improve therapeutic peptides [13,14,15].
While PylRS directly ligates an ncAA onto tRNAPyl, there is no aaRS to form Sec-tRNASec. Rather, Sec is biosynthesized in a tRNA-dependent manner (reviewed in [4]). In bacteria, this first involves the charging of serine (Ser) by seryl-tRNA synthetase (SerRS) to form Ser-tRNASec, followed by the transfer of selenium from selenophosphate by selenocysteine synthase (SelA) for conversion to Sec-tRNASec (Figure 2). In eukaryotes and archaea, Ser-tRNASec is phosphorylated to form O-phosphoseryl-tRNASec (Sep-tRNASec) by Sep-tRNA kinase (PSTK) [16], to which the phosphate group is displaced with selenophosphate by Sep-tRNA:Sec-tRNA synthase (SepSecS) [17,18,19]. Sec-tRNASec delivery to the ribosome is aided by a selenocysteine-specific elongation factor (SelB in bacteria or EFSec in eukaryotes) [20,21]. Furthermore, the Sec insertion sequence (SECIS), an RNA structure in selenoprotein mRNA, recruits the SelB/EFSec-bound Sec-tRNASec to the ribosome for the recoding of a UGA stop codon [22,23] (Figure 2). Given the diverse set of interactions and different mechanisms for Sec incorporation versus PylRS-mediated ncAA incorporation, the task of improving each system requires very different considerations.
When refining Sec- and Pyl-orthogonal translation system (OTS) components for GCE, it is ideal to produce a high amount of the ncAA-tRNA while retaining orthogonality and limiting the effects on cellular fitness. Heterologous aaRS•tRNA pairs for the OTS of a particular host organism are often imported from a different domain of life, since tRNA identity elements and substrate recognition are dissimilar enough to function orthogonally [24]. Moreover, the malleable active site of PylRS allows straightforward directed evolution methods to identify new ncAA-activating variants; however, these variants are polyspecific [25], and mutations that decrease the orthogonality must be selected against. Selenocysteine-OTSs are often used in bacteria or mammalian cells that already have the Sec pathway. Therefore, Sec pathway components are removed to prevent interaction with the OTS. Recent work in Escherichia coli has focused on improving the Sec incorporation efficiency and discovering EF-Tu compatible tRNASec variants for selenoprotein expression without the requirement for SECIS in the coding sequence [26,27,28,29,30,31].
The production of Sec-tRNASec is naturally inefficient compared to canonical aminoacyl-tRNA formation; SerRS serylates tRNASec 100-fold less efficiently than tRNASer [32]. It is likely that this kinetic inefficiency of SerRS correlates with the low demand for Sec incorporation; there are a limited number of proteins requiring Sec. Thus, the most challenging aspect of Sec-OTS engineering is to achieve efficient serylation by SerRS, as well as complete conversion to Sec-tRNASec to ensure limited amounts of Ser misincorporation during selenoprotein expression [28]. Similarly, Pyl-tRNAPyl formation is inefficient compared to other aaRSs and PylRS has a moderate level of catalytic activity [25,33]. It has been a candidate for the evolution of enzyme variants with increased catalytic turnover, as well as more desirable ncAA specificity [34,35].
As a result of increasing ncAA-tRNA concentrations, the cellular levels of the PylRS•tRNAPyl pairs and components of the Sec pathway must be manipulated to out-compete host tRNAs or release factors for the targeted codon, while maintaining cellular fitness. Furthermore, altering the stoichiometry of the Sec-OTS components is important for the efficiency and homogeneity of selenoprotein production [26,29]. Thus, in addition to mutagenesis approaches to improving OTS interactions, the expression levels of each individual component are critical.
Due to their interactions with various parts of the translation machinery, tRNAs are central to achieving highly efficient ncAA incorporation, and both Sec- and Pyl-OTSs can be significantly improved through tRNA engineering. This is often accomplished through rational design, structure-guided mutagenesis, and random mutagenesis. Current molecular biology techniques facilitate the construction of large libraries of mutants, while combining positive and negative selection has been a successful approach to finding better variants. Here, we discuss the aspects of tRNA biology that should be carefully considered prior to OTS engineering and review the recent developments of Sec- and Pyl-OTSs with a main focus on tRNA design.
2. Aspects of Heterologous tRNA Expression
2.1. Identity Elements and Recognition
The identity elements of tRNAs are nucleotides and their modifications, which function as substrate recognition determinants. These determinants are found throughout the tRNA molecule and are essential for interaction with enzymes for aminoacyl-tRNA formation, as well as elongation factors. Moreover, tRNA recognition involves anti-determinant nucleotides and modifications to prevent the binding and charging of non-cognate tRNAs. In some cases, a single nucleotide mutation can change the tRNA identity and allow aminoacylation by a non-cognate aaRS [36]. Similarly, modifications may also confer identity; for example, m1G37 modification of tRNAAsp in yeast is required to inhibit erroneous charging by ArgRS [37]. While some tRNAs (such as tRNAAsp [38]), maintain their identity elements across all domains of life through divergent evolution, domain-specific idiosyncratic features required for aminoacylation are also present [39]. For this reason, aaRS•tRNA pairs can be transplanted from one domain of life to another and function orthogonally with respect to host aminoacylation.
Genetic code expansion designates a particular stop codon, or an “open” codon in genetically recoded organisms, for the insertion of an ncAA. Nonsense suppression is the most common way to insert ncAAs, since recoding is less detrimental to the proteome, given the low occurrence of stop codons. In this regard, the tRNAs of interest for GCE are typically those without identity elements in the anticodon, as their anticodons can be mutated to decode a stop codon of interest, while retaining aminoacylation capabilities. Conversely, if the active site of an aaRS is suitable for engineering ncAA substrate specificity, the anticodon binding domain can be evolved to recognize a nonsense suppressor tRNA [40,41,42,43].
The genetic code naturally expanded to include Sec and Pyl, through the recoding of UGA and UAG, respectively. However, Sec can be efficiently inserted at sense codons [44,45] and improving incorporation in a SECIS-independent manner is achieved through UAG suppression [26,28,29,31]. Anticodon mutations are sufficient to recode sense and stop codons with Sec and Pyl, since cognate SerRS and PylRS do not utilize identity elements in the anticodon loop of tRNASec and tRNAPyl. Thus, ncAA insertion can be easily directed towards a codon of interest using tRNASec and tRNAPyl, within the limitations of the host organism fitness and proteome perturbation.
2.2. Heterologous tRNA Modification and Maturation
Various factors influence the available pools of the aa-tRNA that can be used for peptide synthesis in the cell. These include amino acid and nutrient availability, tRNA expression and maturation (transcription, gene copy number, processing, and modifications), aaRS levels, and tRNA stability and degradation [46]. For GCE applications, the supply of ncAAs is controlled either by adding it in excess amounts to the growth medium or through metabolic engineering of the host organism (e.g., [47]). The biosynthesis and maturation of tRNA are more difficult processes to monitor and control. In E. coli, orthogonal tRNAs can be transcribed from “standard” constitutive and inducible promoters (e.g., lpp, proK, and PBAD). To mimic the coding sequences of bacterial tRNAs, the naturally absent terminal CCA sequence is added to the 3′-end of the archaeal tRNA gene. In contrast, to ensure proper processing in eukaryotes, the 3′-CCA sequence of bacterial orthogonal tRNA genes is typically removed.
While archaeal tRNAs in principle are not orthogonal to eukaryotic aaRSs (one exception being tRNAPyl), bacterial tRNAs are utilized for GCE in eukaryotic hosts [1]. However, the normal transcription of tRNA genes in eukaryotic cells relies on RNA polymerase III, which recognizes A- and B-box promoter elements, present in the tRNA gene itself [48,49]. The majority of prokaryotic tRNAs lack such internal promoter sequences and the engineering of these o-tRNAs may lead to the artificial creation of A- and B-boxes in an o-tRNA variant (see below). To adapt the o-tRNAs of bacterial origin for transcription in yeast, two yeast Pol III promoters—the RPR1 promoter and the SNR52 promoter—have been shown to efficiently drive the expression of E. coli tRNAs [50]. Alternatively, a strong RNA polymerase II promoter with tandem tRNA repeats [51] or the yeast tRNAArg (used as a part of a dicistronic construct) fused upstream of the target tRNA [52] have also been developed.
Between 6.5% and 16.5% of tRNA nucleosides are post-transcriptionally modified, depending on the organisms [53], and over 100 different tRNA modifications have been identified (http://modomics.genesilico.pl/modifications/). Furthermore, tRNA processing is quite complex, sometimes involving intron splicing, trafficking to several subcellular locations [46], and even the ligation of two tRNA halves transcribed from different genes [54]. While the tRNAs used for GCE are orthogonal with respect to endogenous aaRSs, interactions with host modification and processing enzymes is required for function. The addition of tRNA modifications during biosynthesis is important for the stability [55], structure, and function of the molecule [56].
To ensure that an aberrant tRNA is not used for protein synthesis, tRNAs lacking certain modifications are targeted by nucleases for degradation. The nuclear surveillance turnover pathway ensures that a tRNA is properly modified during biosynthesis. For example, yeast pre-tRNAiMet lacking m1A58 is polyadenylated by Trf4, which then triggers nuclease degradation by Rrp6 and the nuclear exosome [57,58]. The modifications m7G and m5C also play a role in tRNA stability. The rapid tRNA decay pathway (RTD) in yeast, involving 5′–3′ exonucleases Rat1 and Xrn1, targets mature tRNA that lack the m7G and m5C modifications [59,60]. These nucleotides provide an additional level of tRNA regulation and can be manipulated (through mutagenesis or the deletion of nonessential tRNA modifying enzymes) to prevent RTD-targeting and increase tRNA abundance, or for targeted degradation to decrease the toxicity of a suppressor tRNA [61].
The modification of tRNA nucleotides also affects codon–anticodon interactions, binding at the ribosomal A site [62], and ultimately the suppression efficiency that is desired for GCE applications. For instance, natural E. coli suppressors depend on the isopentenylation of adenosine 37 for full activity [63,64]. A genetic approach to addressing this issue involves monitoring ncAA incorporation and reporter protein yields across E. coli or yeast strain collections containing deletions and/or the overexpression cassettes of metabolic genes. Recently it was shown that the yield and specificity of O-phosphoserine incorporation is significantly improved by the deletion of cysteine desulfurase and the overexpression of E. coli dimethylallyltransferase (MiaA) and pseudouridine synthase (TruB) [65]. Furthermore, a yeast study involving the removal of modifications by single gene deletions from U34, U35, A37, U47 and C48 in the anticodon stem-loop impairs nonsense suppression, with the strongest effect observed for U34 and A37. Interestingly, the overexpression of eEF1a rescues the activity of an ochre suppressor tRNA (SUP4) and other non-suppressor tRNAs that lack modifications [66]. Thus, when designing suppressor tRNAs for GCE, tRNA modifications must be maintained or compensated for, such that tRNA stability and ncAA incorporation is not compromised.
3. When Amino Acid Biosynthesis is o-tRNA-Dependent: Challenges in tRNASec Engineering
The biosynthesis of Sec-tRNASec and its delivery to the ribosome is complex compared to the canonical amino acid pathway and involves several interactions with different portions of tRNASec. The major challenge in engineering tRNASec for the more efficient incorporation of Sec is to improve serylation, while also having complete conversion of Ser-tRNASec to Sec-tRNASec. In addition to this, the requirement of a SECIS sequence directly after UGA necessitates an EF-Tu-mediated Sec insertion pathway for the design and expression of selenoproteins in bacteria.
3.1. tRNASec Interactions
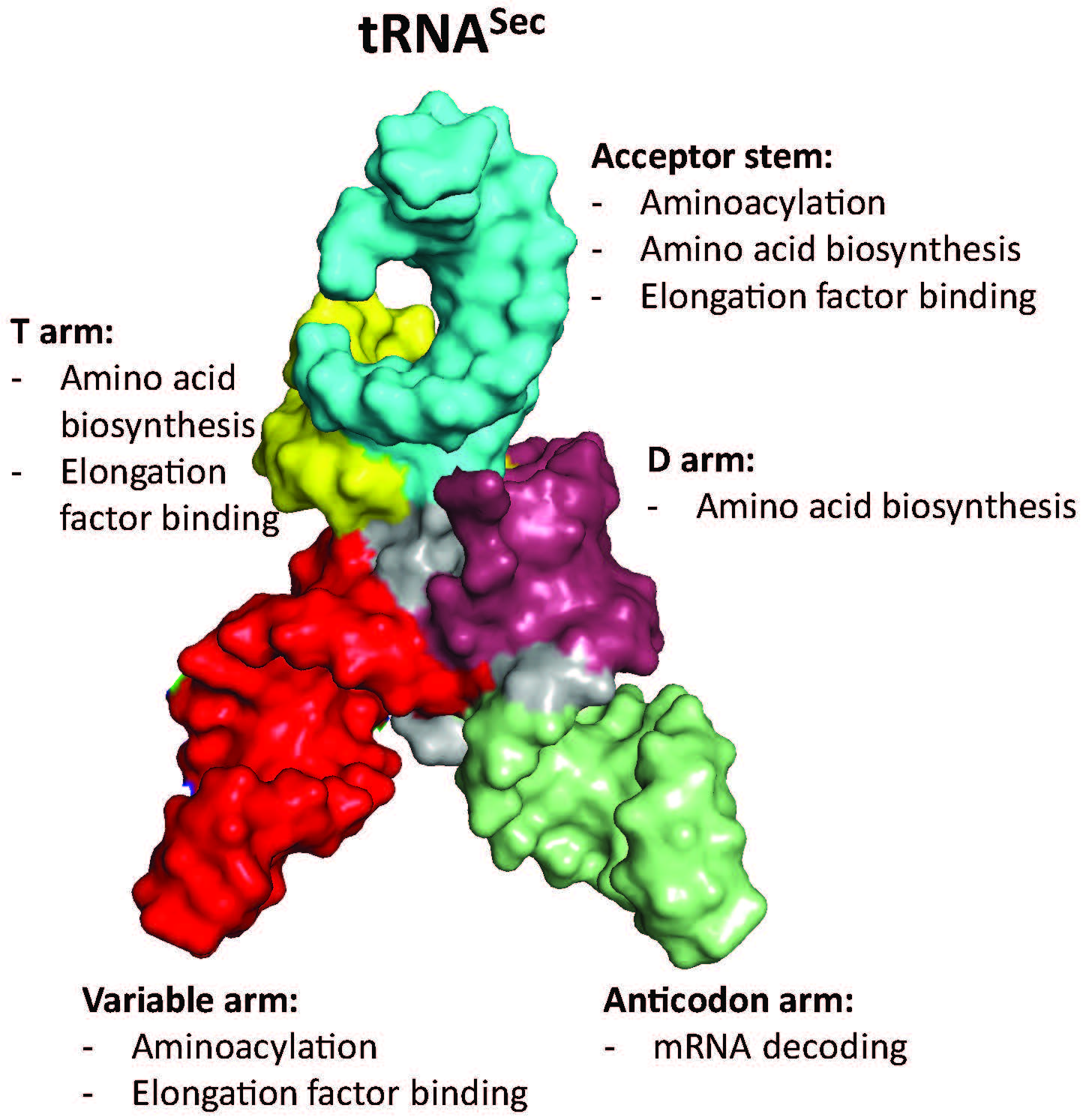
The first step in Sec biosynthesis is the charging of tRNASec with Ser by SerRS (Figure 2). SerRS lacks an anticodon binding domain, and changes to the anticodon stem-loop do not affect aminoacylation [67]. Rather, SerRS recognizes a long variable arm, a G73 discriminator base, and identity elements in the acceptor and D stems [68,69,70,71,72], which are conserved between tRNASer and tRNASec (Figure 3). These elements contribute to the structural features and shape of the tRNA and are important for the backbone and sequence-specific interactions for recognition by SerRS [68]. Of these features, the variable arm is most critical for aminoacylation. SerRS possess an N-terminal helical extension that interacts with the variable arm of tRNASer and tRNASec, and properly orients the tRNA 3′ end for aminoacylation [68,73,74]. The overall length of the variable arm is more important than the sequence; the insertion of only one or two nucleotides in the variable arm of tRNALeu and tRNATyr, respectively, confers serylation activity and the deletion of a single base pair from the tRNASec variable arm improves serylation 2–3 fold [32,70,75]. It is therefore not surprising that the variable arm accounts for the largest influence on the Km/kcat of aminoacylation [67].
Identity elements of the tRNASec extend beyond aminoacylation and include features of SelA and SelB interactions. Whereas canonical tRNAs have a 12-base-pair amino acid acceptor branch (7/5; consisting of a seven-base-pair acceptor stem and a five-base-pair T stem) that is recognized by EF-Tu/eEF1a, tRNASec has a longer 13-base-pair acceptor branch (8/5 or 9/4). The deletion of a base pair from the acceptor stem of E. coli tRNASec to resemble that of canonical tRNASer abolishes UGA read-through with Sec [45], likely due to the disruption of the complex formation of tRNASec with SelA and SelB [32]. In addition to the effects of the acceptor stem length on SelA recognition, nucleotides in the D arm form a unique structure compared to tRNASer, which is the basis of SelA-tRNASec interaction [76].
Comparisons of SelB and EF-Tu complex structures show similarities of acceptor stem binding, but also unique domains and motifs that provide tRNA specificity. The N-terminal half of SelB consists of three domains, named D1, D2, and D3, that are analogous to those of EF-Tu [77]. D1 makes up the GTP-binding domain whereas, D2 and D3 consist of β-barrel-like and β-barrel structures for tRNA binding. Unique to SelB is a fourth domain (D4) comprised of four wing-helix motifs that recruit SelB to SECIS [78]. The structures of the SelB-Sec-tRNASec complex obtained from single-particle cryo-electron microscopy depict how the linker region between D3 and D4 binds and distorts the variable arm of tRNASec, while an extended loop of D3 interacts with the acceptor and T stems [79]. In conjunction with the positively-charged SelB binding pocket, which provides affinity for the selenol group of Sec, and the altered variable arm orientation of tRNASec compared to tRNASer, D3 and the linker between D3 and D4 of SelB provide Sec-tRNASec specificity.
3.2. Converting tRNASec Recognition from SelB to EF-Tu
The acceptor stem of the tRNASec posed a challenge for engineering the EF-Tu-mediated Sec insertion. Although the binding specificity of tRNASec can be switched from SelB to EF-Tu by shortening the acceptor stem [32], the eight-base-pair stem is important for the interaction with SelA. However, three base pairs in the T stem (49:65, 50:64, and 51:63) modulate the binding affinity of EF-Tu in a sequence-dependent manner [80]. In the same region, tRNASec has different bases. Moreover, the last base pair of the acceptor stem and the first two base pairs of the T stem of tRNASec are anti-determinants of EF-Tu complexed with GTP [81].
The first generation tRNASec for EF-Tu recognition, named tRNAUTu (U for Sec and Tu for EF-Tu), was designed using E. coli tRNASer as a scaffold with the first seven base pairs of the E. coli tRNASec acceptor stem [31]. The last base pair of the tRNAUTu acceptor stem was transplanted from tRNASer to eliminate the EF-Tu anti-determinant position. Serylation of tRNAUTu was as efficient as canonical tRNASer, however, the Ser to Sec conversion was hampered, which led to ~30% Ser misincorporation. Nonetheless, tRNAUTu was successfully used to site-specifically incorporate Sec into selenoproteins of bacterial and human origin in a SECIS-independent manner.
3.3. Improving Ser-to-Sec Conversion
Complementary approaches were taken to address the incomplete conversion of Ser-tRNAUTu to Sec-tRNAUTu. E. coli tRNASec was used as a scaffold for the random mutagenesis of the EF-Tu anti-determinant base pairs C7:G66, G49:U65, and C50:G64. A Sec-specific NMC-A β-lactamase reporter was selected as an efficient tRNASec suppressor containing G7:C66, U49:G65, and C50:U64, which was named tRNASecUX [29]. In order to achieve nearly complete conversion of Ser to Sec, SelA expression was elevated, the tRNASecUX dosage was decreased, and PSTK was co-expressed to form a Sep-tRNASecUX intermediate, which would remain a substrate for SelA but not for EF-Tu prior to Sec conversion.
Other studies have built on tRNAUTu to improve Ser to Sec conversion. Using the structure of Aquifex aeolicus SelA in a complex with Thermus tengcongensis tRNASec, twenty-nine different tRNAUTu variants were rationally designed to include tRNASec features that interact with SelA, while maintaining those that are required for EF-Tu binding. E. coli FDHH was used as a Sec insertion reporter in a sensitive colorimetric assay to identify the best variant, named tRNAUTuX, which differed from tRNAUTu at 11 positions [28]. Kinetic assays confirmed that the serylation of tRNAUTuX was comparable to tRNAUTu and tRNASec. Ser-to-Sec conversion was increased to 90%, reaching a similar conversion rate as E. coli tRNASec. Furthermore, Fourier transform ion cyclotron resonance (FT-ICR) mass spectrometry analysis confirmed Sec insertion by tRNAUTuX into the selenoprotein, Grx1, but did not detect a peak corresponding to Ser insertion. More recently, tRNAUTu was used as a template for the generation of chimeric molecules to improve Sec incorporation and selenoprotein yields. It was found that a single base change of A59C in tRNAUTu, generating a molecule named tRNAUTu6, resulted in the highest expression levels of human GPx1 and nearly 90% Sec incorporation [27].
3.4. Different tRNASec Structures for the Optimization of Selenoprotein Production
In a bioinformatic search for novel tRNASec molecules, a group of tRNAs with unusual cloverleaf structures were identified, named allo-tRNA [82,83]. Certain allo-tRNA species had tRNASer identities and functioned as efficient amber suppressors with Ser [82]. Allo-tRNAs also contain SelA identity elements, but have a 12-base-pair acceptor branch as opposed to the 13-pair branch present in most tRNASec molecules. SelA from Aeromonas salmonicida subsp. pectinolytica 34mel (As) was coupled with allo-tRNA for selenoprotein expression, since its cognate tRNASec also possesses a 12-base-pair acceptor branch [26]. Allo-tRNA nucleotides in the D stem and acceptor stem were mutated to include As tRNASec identities. In addition, the stoichiometry of allo-tRNA to As SelA was altered to ensure the complete sequestration of the tRNA for Ser-to-Sec conversion while also maintaining non-toxic levels of As SelA. Further optimizations and metabolic engineering efforts created a Sec-OTS consisting of allo-tRNAUTu2D, As SelD, As SelA, and Treponema denticola Trx1. Along with the high selenoprotein yields obtained with a Sec incorporation efficiency estimated at >90%, the stand-alone capabilities of this system make it ideal for use in other organisms [26].
4. Absolutely Orthogonal? Unique Features of tRNAPyl
Compared to the Sec system, the use of Pyl-OTS is comparatively less challenging, as its tRNA is orthogonal in the majority of model organisms used for GCE [84]; the enzyme is also orthogonal to both cellular tRNAs, as well as natural/canonical AAs [85]. Both bacterial and eukaryotic elongation factors accept tRNAPyl, and the AA-binding pocket can be separately adapted to accept some bulkier ncAAs [86]. Attempts to advance ncAA delivery by tRNAPyl engineering include those aiming to improve its compatibility with the cellular machinery of the host. In E. coli, tRNAPyl was evolved by targeting the EF-Tu-binding regions [87], although the optimizing mutations present in tRNAPylOPT may be more suitable for the delivery of one particular ncAA and less for the other (e.g., Nε-acetyllyine vs. 3-cyano-phenylalanine) [88]. The need to separately evolve an o-tRNA for a variety of “cognate” ncAAs or a variety of anticodons may require tunable binding by EF-Tu and the ribosome; while the stability of the EF-Tu•ncAA-tRNA complex reflects additive contributions by the ncAA and T-stem base pairs of the o-tRNA [80,89], the strength of codon–anticodon binding correlates with the nucleotide composition of the tRNA core [90]. For efficient expression in mammalian systems, a stabilizing mutation in the anticodon stem has been used (U29aC, Figure 4) [91,92]. By introducing elements conserved in human tRNAs, a better performing tRNAPyl was evolved. Mutations in the D-stem, D-loop, T-loop and the anticodon-stem U29aC proved to be indispensable for high activity [93]; compared to wild type tRNAPyl, the use of this variant in HEK293 cells improved the incorporation of two ncAAs, Nε-carbobenzyloxy-lysine (Z-lysine) and Nε-(tert-butoxycarbonyl)-lysine (Boc-lysine). Interestingly, a chimera between mitochondrial (mt) tRNASer and Methanosarcina mazei tRNAPyl improved the insertion of Boc-lysine selectively (C15) [93]. Earlier attempts at using mttRNASer in E. coli failed, due to the lack of orthogonality [94]. The improved activity of M15 and C15 variants in mammalian cell lines may have to do with the appearance of the B-box in the T-arm of the variants; prokaryotic o-tRNAs are usually placed under the external promoter, such as U6, but the endogenous tRNAs are transcribed from internal A- and B-box promoters [48].
One of the distinct features of the Pyl system is the minimal variable loop of tRNAPyl, which together with the T-loop forms a dipped surface [35,95] (Figure 4). From the crystal structure of the N-terminal domain of M. mazei PylRS it is evident that this minimalistic variable loop is a prerequisite for effective binding, as a larger variable loop would sterically clash with the N-domain [35]. In addition to M. mazei, Desulfitobacterium hafniense Pyl-OTS was employed in E. coli, either with its original N-terminal domain, or as a fusion with the recombinant (chimeric) N-domain of the archaeal system [96]. However, this system is not functional in mammalian cells [93]. As the N-terminal domain binds tRNAPyl with extremely high affinity [4], this element is likely to be an important contributor to (almost universal) Pyl-OTS orthogonality.
However, some organisms do not possess an equivalent to this N-domain [99], suggesting an alternative mode of recognition. This fact was recently exploited to develop mutually orthogonal Pyl-OTSs in E. coli [98] and mammalian cell lines [97,100]. Two PylRS enzymes that utilize the C-domain only (Methanomethylophilus alvus and methanogenic archaeon ISO4-G1) are highly active in E. coli [98]. Their cognate tRNAs retain some characteristic MmtRNAPyl features (such as the identity of the discriminator base G73, or the minimalistic D-loop) but also diverge in the nucleotide composition of the acceptor stem and in the probable structure of the anticodon stem (Figure 4). Given that the M. mazei and M. alvus/G1 systems are not fully orthogonal, rational engineering was employed in order to generate MatRNAPyl that would be recognized by MaPylRS and not MmPylRS. Variation of the nucleotide composition of the variable arm and/or its length allowed the generation of successful MatRNAPyl variants. Given the malleability of the PylRS active site, orthogonality to other OTSs [26,101], together with high activity of Pyl-OTSs in the bacteria and cells of higher eukaryotes [6], it is foreseeable that this dual encoding system will be commonly used.
The creation of multiple, mutually orthogonal OTSs is inherently related to the number of liberated codons that can be targeted for ncAA incorporation. In addition to UAG-directed incorporation, Pyl-OTS was also employed for ncAA incorporation in response to rare arginine (AGG) codons in E. coli, alone [102] or in tandem with Methanocaldococcus jannaschii Tyr-OTS [26]. A similar strategy was attempted in Mycoplasma capricolum, which possesses only six arginine CGG codons that should, in theory, facilitate the reassignment (Arg-to-Pyl) [103]. However, upon mutation of the tRNAPyl anticodon to CCG this almost universally orthogonal tRNA becomes a substrate for endogenous ArgRS. In conclusion, while the anticodon-blind recognition of PylRS allows the anticodon of tRNAPyl to be mutated into any nucleotide triplet, synonymous anticodons (such as CCU and CCG) can be recognized by host aaRSs with very different affinity, causing one tRNAPyl variant to lose its initial orthogonality.
5. Conclusions/Outlook
Improvements to OTSs have been emerging rapidly in recent years and are valuable for the accurate and efficient production of proteins containing ncAAs. The increasing amount of sequence data and bioinformatic/structural analyses reveal new molecules and novel mechanisms that help enhance each system. Moreover, advanced molecular cloning and directed evolution techniques help further shape the molecules that nature has provided into molecules that are better suited for the incorporation of ncAAs. tRNAs interact with each component of an OTS in the process of bringing the ncAA to the ribosome to insert a particular ncAA during peptide synthesis. For this reason, finding the best tRNA variant is critical for OTS developments. Our expanding knowledge of tRNA processing, maturation, and interaction mechanisms has guided tRNA engineering towards this goal. As we continue to learn more from nature and as technologies advance, it is conceivable that peptides with unique properties will be produced with significant industrial and medical implications.
Author Contributions
Conceptualization, K.S.H., A.C., and D.S.; Writing-Original Draft Preparation, K.S.H., A.C., and D.S.; Writing-Review and Editing, K.S.H., A.C., and D.S.; Visualization, K.S.H. and A.C.; Supervision, D.S.; Funding Acquisition, D.S.
Funding
Research in the Söll laboratory is supported by grants from the US National Institutes of Health (NIH) (R35GM122560), the US National Science Foundation (CHE-1740549) and from the Division of Chemical Sciences, Geosciences, and Biosciences, Office of Basic Energy Sciences of the Department of Energy (DE-FG02-98ER20311) to D.S.
Acknowledgments
We thank Oscar Vargas-Rodriguez, Hui Si Kwok, and Jeffery Tharp for critical discussion during the preparation of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chin, J.W. Expanding and reprogramming the genetic code of cells and animals. Annu. Rev. Biochem. 2014, 83, 379–408. [Google Scholar] [CrossRef] [PubMed]
- Mukai, T.; Lajoie, M.J.; Englert, M.; Soll, D. Rewriting the Genetic Code. Annu. Rev. Microbiol. 2017, 71, 557–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, D.D.; Schultz, P.G. Playing with the molecules of life. ACS Chem. Biol. 2018, 13, 854–870. [Google Scholar] [CrossRef] [PubMed]
- Herring, S.; Ambrogelly, A.; Gundllapalli, S.; O’Donoghue, P.; Polycarpo, C.R.; Söll, D. The amino-terminal domain of pyrrolysyl-tRNA synthetase is dispensable in vitro but required for in vivo activity. FEBS Lett. 2007, 581, 3197–3203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, R.; Krzycki, J.A. PylSn and the homologous N-terminal domain of pyrrolysyl-tRNA synthetase bind the tRNA that is essential for the genetic encoding of pyrrolysine. J. Biol. Chem. 2012, 287, 32738–32746. [Google Scholar] [CrossRef] [PubMed]
- Wan, W.; Tharp, J.M.; Liu, W.R. Pyrrolysyl-tRNA synthetase: An ordinary enzyme but an outstanding genetic code expansion tool. Biochim. Biophys. Acta 2014, 1844, 1059–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, J.W.; Cropp, T.A.; Anderson, J.C.; Mukherji, M.; Zhang, Z.; Schultz, P.G. An expanded eukaryotic genetic code. Science 2003, 301, 964–967. [Google Scholar] [CrossRef] [PubMed]
- Greiss, S.; Chin, J.W. Expanding the genetic code of an animal. J. Am. Chem. Soc. 2011, 133, 14196–14199. [Google Scholar] [CrossRef] [PubMed]
- Si, L.; Xu, H.; Zhou, X.; Zhang, Z.; Tian, Z.; Wang, Y.; Wu, Y.; Zhang, B.; Niu, Z.; Zhang, C.; et al. Generation of influenza A viruses as live but replication-incompetent virus vaccines. Science 2016, 354, 1170–1173. [Google Scholar] [CrossRef] [PubMed]
- Bianco, A.; Townsley, F.M.; Greiss, S.; Lang, K.; Chin, J.W. Expanding the genetic code of Drosophila melanogaster. Nat. Chem. Biol. 2012, 8, 748–750. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.C.; Schultz, P.G. Adding new chemistries to the genetic code. Annu. Rev. Biochem. 2010, 79, 413–444. [Google Scholar] [CrossRef] [PubMed]
- Reich, H.J.; Hondal, R.J. Why nature chose selenium. ACS Chem. Biol. 2016, 11, 821–841. [Google Scholar] [CrossRef] [PubMed]
- Arai, K.; Takei, T.; Okumura, M.; Watanabe, S.; Amagai, Y.; Asahina, Y.; Moroder, L.; Hojo, H.; Inaba, K.; Iwaoka, M. Preparation of Selenoinsulin as a Long-Lasting Insulin Analogue. Angew. Chem. Int. Ed. Engl. 2017, 56, 5522–5526. [Google Scholar] [CrossRef] [PubMed]
- Metanis, N.; Hilvert, D. Natural and synthetic selenoproteins. Curr. Opin. Chem. Biol. 2014, 22, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Shchedrina, V.A.; Novoselov, S.V.; Malinouski, M.Y.; Gladyshev, V.N. Identification and characterization of a selenoprotein family containing a diselenide bond in a redox motif. Proc. Natl. Acad. Sci. USA 2007, 104, 13919–13924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, B.A.; Xu, X.M.; Kryukov, G.V.; Rao, M.; Berry, M.J.; Gladyshev, V.N.; Hatfield, D.L. Identification and characterization of phosphoseryl-tRNA[Ser]Sec kinase. Proc. Natl. Acad. Sci. USA 2004, 101, 12848–12853. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.M.; Carlson, B.A.; Mix, H.; Zhang, Y.; Saira, K.; Glass, R.S.; Berry, M.J.; Gladyshev, V.N.; Hatfield, D.L. Biosynthesis of selenocysteine on its tRNA in eukaryotes. PLoS Biol. 2007, 5, e4. [Google Scholar] [CrossRef] [PubMed]
- Palioura, S.; Sherrer, R.L.; Steitz, T.A.; Söll, D.; Simonovic, M. The human SepSecS-tRNASec complex reveals the mechanism of selenocysteine formation. Science 2009, 325, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Palioura, S.; Salazar, J.C.; Su, D.; O’Donoghue, P.; Hohn, M.J.; Cardoso, A.M.; Whitman, W.B.; Soll, D. RNA-dependent conversion of phosphoserine forms selenocysteine in eukaryotes and archaea. Proc. Natl. Acad. Sci. USA 2006, 103, 18923–18927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forchhammer, K.; Leinfelder, W.; Böck, A. Identification of a novel translation factor necessary for the incorporation of selenocysteine into protein. Nature 1989, 342, 453–456. [Google Scholar] [CrossRef] [PubMed]
- Fagegaltier, D.; Hubert, N.; Yamada, K.; Mizutani, T.; Carbon, P.; Krol, A. Characterization of mSelB, a novel mammalian elongation factor for selenoprotein translation. EMBO J. 2000, 19, 4796–4805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Squires, J.E.; Berry, M.J. Eukaryotic selenoprotein synthesis: Mechanistic insight incorporating new factors and new functions for old factors. IUBMB Life 2008, 60, 232–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donovan, J.; Copeland, P.R. Threading the needle: Getting selenocysteine into proteins. Antioxid. Redox Signal. 2010, 12, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Giege, R.; Sissler, M.; Florentz, C. Universal rules and idiosyncratic features in tRNA identity. Nucleic Acids Res. 1998, 26, 5017–5035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.T.; Wang, Y.S.; Nakamura, A.; Eiler, D.; Kavran, J.M.; Wong, M.; Kiessling, L.L.; Steitz, T.A.; O’Donoghue, P.; Söll, D. Polyspecific pyrrolysyl-tRNA synthetases from directed evolution. Proc. Natl. Acad. Sci. USA 2014, 111, 16724–16729. [Google Scholar] [CrossRef] [PubMed]
- Proc Natl Acad Sci U S AFEBS LettOhtake, K.; Mukai, T.; Iraha, F.; Takahashi, M.; Haruna, K.-i.; Date, M.; Yokoyama, K.; Sakamoto, K. Engineering an automaturing transglutaminase with enhanced thermostability by genetic code expansion with two codon reassignments. ACS Synth. Biol. 2018, 7, 2170–2176. [Google Scholar] [CrossRef]
- Fan, Z.; Song, J.; Guan, T.; Lv, X.; Wei, J. Efficient expression of glutathione peroxidase with chimeric tRNA in amber-less Escherichia coli. ACS Synth. Biol. 2018, 7, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.; Bröcker, M.J.; Prat, L.; Ip, K.; Chirathivat, N.; Feiock, A.; Veszpremi, M.; Söll, D. A synthetic tRNA for EF-Tu mediated selenocysteine incorporation in vivo and in vitro. FEBS Lett. 2015, 589, 2194–2199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thyer, R.; Robotham, S.A.; Brodbelt, J.S.; Ellington, A.D. Evolving tRNASec for efficient canonical incorporation of selenocysteine. J. Am. Chem. Soc. 2015, 137, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Haruna, K.; Alkazemi, M.H.; Liu, Y.; Söll, D.; Englert, M. Engineering the elongation factor Tu for efficient selenoprotein synthesis. Nucleic Acids Res. 2014, 42, 9976–9983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldag, C.; Bröcker, M.J.; Hohn, M.J.; Prat, L.; Hammond, G.; Plummer, A.; Söll, D. Rewiring translation for elongation factor Tu-dependent selenocysteine incorporation. Angew. Chem. Int. Ed. Engl. 2013, 52, 1441–1445. [Google Scholar] [CrossRef] [PubMed]
- Baron, C.; Böck, A. The length of the aminoacyl-acceptor stem of the selenocysteine-specific tRNASec of Escherichia coli is the determinant for binding to elongation factors SELB or Tu. J. Biol. Chem. 1991, 266, 20375–20379. [Google Scholar] [PubMed]
- Li, W.T.; Mahapatra, A.; Longstaff, D.G.; Bechtel, J.; Zhao, G.; Kang, P.T.; Chan, M.K.; Krzycki, J.A. Specificity of pyrrolysyl-tRNA synthetase for pyrrolysine and pyrrolysine analogs. J. Mol. Biol. 2009, 385, 1156–1164. [Google Scholar] [CrossRef] [PubMed]
- Bryson, D.I.; Fan, C.; Guo, L.T.; Miller, C.; Söll, D.; Liu, D.R. Continuous directed evolution of aminoacyl-tRNA synthetases. Nat. Chem. Biol. 2017, 13, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Miller, C.; Guo, L.T.; Ho, J.M.L.; Bryson, D.I.; Wang, Y.S.; Liu, D.R.; Söll, D. Crystal structures reveal an elusive functional domain of pyrrolysyl-tRNA synthetase. Nat. Chem. Biol. 2017, 13, 1261–1266. [Google Scholar] [CrossRef] [PubMed]
- Auld, D.S.; Schimmel, P. Switching recognition of two tRNA synthetases with an amino acid swap in a designed peptide. Science 1995, 267, 1994–1996. [Google Scholar] [CrossRef] [PubMed]
- Perret, V.; Garcia, A.; Grosjean, H.; Ebel, J.P.; Florentz, C.; Giege, R. Relaxation of a transfer RNA specificity by removal of modified nucleotides. Nature 1990, 344, 787–789. [Google Scholar] [CrossRef] [PubMed]
- Becker, H.D.; Giege, R.; Kern, D. Identity of prokaryotic and eukaryotic tRNAAsp for aminoacylation by aspartyl-tRNA synthetase from Thermus thermophilus. Biochemistry 1996, 35, 7447–7458. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.P.; RajBhandary, U.L. Mutants of Escherichia coli initiator tRNA that suppress amber codons in Saccharomyces cerevisiae and are aminoacylated with tyrosine by yeast extracts. Proc. Natl. Acad. Sci. USA 1991, 88, 11378–11382. [Google Scholar] [CrossRef] [PubMed]
- Gan, R.; Perez, J.G.; Carlson, E.D.; Ntai, I.; Isaacs, F.J.; Kelleher, N.L.; Jewett, M.C. Translation system engineering in Escherichia coli enhances non-canonical amino acid incorporation into proteins. Biotechnol. Bioeng. 2017, 114, 1074–1086. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Oh, S.; Yang, A.; Kim, J.; Soll, D.; Lee, D.; Park, H.S. A facile strategy for selective incorporation of phosphoserine into histones. Angew. Chem. Int. Ed. Engl. 2013, 52, 5771–5775. [Google Scholar] [CrossRef] [PubMed]
- Rogerson, D.T.; Sachdeva, A.; Wang, K.; Haq, T.; Kazlauskaite, A.; Hancock, S.M.; Huguenin-Dezot, N.; Muqit, M.M.; Fry, A.M.; Bayliss, R.; et al. Efficient genetic encoding of phosphoserine and its nonhydrolyzable analog. Nat. Chem. Biol. 2015, 11, 496–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, A.; Xiao, H.; Schultz, P.G. Evolution of multiple, mutually orthogonal prolyl-tRNA synthetase/tRNA pairs for unnatural amino acid mutagenesis in Escherichia coli. Proc. Natl. Acad. Sci. USA 2012, 109, 14841–14846. [Google Scholar] [CrossRef] [PubMed]
- Bröcker, M.J.; Ho, J.M.; Church, G.M.; Söll, D.; O’Donoghue, P. Recoding the genetic code with selenocysteine. Angew. Chem. Int. Ed. Engl. 2014, 53, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Baron, C.; Heider, J.; Böck, A. Mutagenesis of selC, the gene for the selenocysteine-inserting tRNA-species in E. coli: Effects on in vivo function. Nucleic Acids Res. 1990, 18, 6761–6766. [Google Scholar] [CrossRef] [PubMed]
- Phizicky, E.M.; Hopper, A.K. tRNA biology charges to the front. Genes Dev. 2010, 24, 1832–1860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinfeld, J.B.; Aerni, H.R.; Rogulina, S.; Liu, Y.; Rinehart, J. Expanded cellular amino acid pools containing phosphoserine, phosphothreonine, and phosphotyrosine. ACS Chem. Biol. 2014, 9, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Galli, G.; Hofstetter, H.; Birnstiel, M.L. Two conserved sequence blocks within eukaryotic tRNA genes are major promoter elements. Nature 1981, 294, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Sharp, S.; DeFranco, D.; Dingermann, T.; Farrell, P.; Soll, D. Internal control regions for transcription of eukaryotic tRNA genes. Proc. Natl. Acad. Sci. USA 1981, 78, 6657–6661. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, L. Genetic incorporation of unnatural amino acids into proteins in yeast. Methods Mol. Biol. 2012, 794, 199–213. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Schultz, P.G.; Brock, A. An improved system for the generation and analysis of mutant proteins containing unnatural amino acids in Saccharomyces cerevisiae. J. Mol. Biol. 2007, 371, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Otter, C.A.; Straby, K.B. Transcription of eukaryotic genes with impaired internal promoters: The use of a yeast tRNA gene as promoter. J. Biotechnol. 1991, 21, 289–293. [Google Scholar] [CrossRef]
- Machnicka, M.A.; Olchowik, A.; Grosjean, H.; Bujnicki, J.M. Distribution and frequencies of post-transcriptional modifications in tRNAs. RNA Biol. 2014, 11, 1619–1629. [Google Scholar] [CrossRef] [PubMed]
- Randau, L.; Munch, R.; Hohn, M.J.; Jahn, D.; Söll, D. Nanoarchaeum equitans creates functional tRNAs from separate genes for their 5′- and 3′-halves. Nature 2005, 433, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, C.; Lünse, E.C.; Mörl, M. tRNA Modifications: Impact on Structure and Thermal Adaptation. Biomolecules 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Väre, Y.V.; Eruysal, R.E.; Narendran, A.; Sarachan, L.K.; Agris, F.P. Chemical and conformational diversity of modified nucleosides affects trna structure and function. Biomolecules 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Kadaba, S.; Wang, X.; Anderson, J.T. Nuclear RNA surveillance in Saccharomyces cerevisiae: Trf4p-dependent polyadenylation of nascent hypomethylated tRNA and an aberrant form of 5S rRNA. RNA 2006, 12, 508–521. [Google Scholar] [CrossRef] [PubMed]
- Kadaba, S.; Krueger, A.; Trice, T.; Krecic, A.M.; Hinnebusch, A.G.; Anderson, J. Nuclear surveillance and degradation of hypomodified initiator tRNAMet in S. cerevisiae. Genes Dev. 2004, 18, 1227–1240. [Google Scholar] [CrossRef] [PubMed]
- Dewe, J.M.; Whipple, J.M.; Chernyakov, I.; Jaramillo, L.N.; Phizicky, E.M. The yeast rapid tRNA decay pathway competes with elongation factor 1A for substrate tRNAs and acts on tRNAs lacking one or more of several modifications. RNA 2012, 18, 1886–1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexandrov, A.; Chernyakov, I.; Gu, W.; Hiley, S.L.; Hughes, T.R.; Grayhack, E.J.; Phizicky, E.M. Rapid tRNA decay can result from lack of nonessential modifications. Mol. Cell 2006, 21, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Berg, M.D.; Hoffman, K.S.; Genereaux, J.; Mian, S.; Trussler, R.S.; Haniford, D.B.; O’Donoghue, P.; Brandl, C.J. Evolving mistranslating tRNAs through a phenotypically ambivalent intermediate in Saccharomyces cerevisiae. Genetics 2017, 206, 1865–1879. [Google Scholar] [CrossRef] [PubMed]
- Grosjean, H.; Westhof, E. An integrated, structure- and energy-based view of the genetic code. Nucleic Acids Res. 2016, 44, 8020–8040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gefter, M.L.; Russell, R.L. Role modifications in tyrosine transfer RNA: A modified base affecting ribosome binding. J. Mol. Biol. 1969, 39, 145–157. [Google Scholar] [CrossRef]
- Laten, H.; Gorman, J.; Böck, R.M. Isopentenyladenosine deficient tRNA from an antisuppressor mutant of Saccharomyces cerevisiae. Nucleic Acids Res. 1978, 5, 4329–4342. [Google Scholar] [CrossRef] [PubMed]
- Crnkovic, A.; Vargas-Rodriguez, O.; Merkuryev, A.; Söll, D. Effects of heterologous tRNA modifications on the production of proteins containing noncanonical amino acids. Bioengineering 2018, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- Klassen, R.; Schaffrath, R. Collaboration of tRNA modifications and elongation factor eEF1A in decoding and nonsense suppression. Sci. Rep. 2018, 8, 12749. [Google Scholar] [CrossRef] [PubMed]
- Sampson, J.R.; Saks, M.E. Contributions of discrete tRNASer domains to aminoacylation by E. coli seryl-tRNA synthetase: A kinetic analysis using model RNA substrates. Nucleic Acids Res. 1993, 21, 4467–4475. [Google Scholar] [CrossRef] [PubMed]
- Biou, V.; Yaremchuk, A.; Tukalo, M.; Cusack, S. The 2.9 Å crystal structure of T. thermophilus seryl-tRNA synthetase complexed with tRNASer. Science 1994, 263, 1404–1410. [Google Scholar] [CrossRef] [PubMed]
- Normanly, J.; Ollick, T.; Abelson, J. Eight base changes are sufficient to convert a leucine-inserting tRNA into a serine-inserting tRNA. Proc. Natl. Acad. Sci. USA 1992, 89, 5680–5684. [Google Scholar] [CrossRef] [PubMed]
- Himeno, H.; Hasegawa, T.; Ueda, T.; Watanabe, K.; Shimizu, M. Conversion of aminoacylation specificity from tRNATyr to tRNASer in vitro. Nucleic Acids Res. 1990, 18, 6815–6819. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.J.; Söll, D. Discrimination between glutaminyl-tRNA synthetase and seryl-tRNA synthetase involves nucleotides in the acceptor helix of tRNA. Proc. Natl. Acad. Sci. USA 1988, 85, 6627–6631. [Google Scholar] [CrossRef] [PubMed]
- Normanly, J.; Ogden, R.C.; Horvath, S.J.; Abelson, J. Changing the identity of a transfer RNA. Nature 1986, 321, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Itoh, Y.; Sekine, S.; Suetsugu, S.; Yokoyama, S. Tertiary structure of bacterial selenocysteine tRNA. Nucleic Acids Res. 2013, 41, 6729–6738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Guo, Y.; Tian, Q.; Jia, Q.; Gao, Y.; Zhang, Q.; Zhou, C.; Xie, W. SerRS-tRNASec complex structures reveal mechanism of the first step in selenocysteine biosynthesis. Nucleic Acids Res. 2015, 43, 10534–10545. [Google Scholar] [CrossRef] [PubMed]
- Himeno, H.; Yoshida, S.; Soma, A.; Nishikawa, K. Only one nucleotide insertion to the long variable arm confers an efficient serine acceptor activity upon Saccharomyces cerevisiae tRNALeu in vitro. J. Mol. Biol. 1997, 268, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Itoh, Y.; Bröcker, M.J.; Sekine, S.; Hammond, G.; Suetsugu, S.; Söll, D.; Yokoyama, S. Decameric SelA•tRNASec ring structure reveals mechanism of bacterial selenocysteine formation. Science 2013, 340, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Selmer, M.; Su, X.D. Crystal structure of an mRNA-binding fragment of Moorella thermoacetica elongation factor SelB. EMBO J. 2002, 21, 4145–4153. [Google Scholar] [CrossRef] [PubMed]
- Kromayer, M.; Wilting, R.; Tormay, P.; Böck, A. Domain structure of the prokaryotic selenocysteine-specific elongation factor SelB. J. Mol. Biol. 1996, 262, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Fischer, N.; Neumann, P.; Bock, L.V.; Maracci, C.; Wang, Z.; Paleskava, A.; Konevega, A.L.; Schroder, G.F.; Grubmuller, H.; Ficner, R.; et al. The pathway to GTPase activation of elongation factor SelB on the ribosome. Nature 2016, 540, 80–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrader, J.M.; Chapman, S.J.; Uhlenbeck, O.C. Understanding the sequence specificity of tRNA binding to elongation factor Tu using tRNA mutagenesis. J. Mol. Biol. 2009, 386, 1255–1264. [Google Scholar] [CrossRef] [PubMed]
- Rudinger, J.; Hillenbrandt, R.; Sprinzl, M.; Giege, R. Antideterminants present in minihelixSec hinder its recognition by prokaryotic elongation factor Tu. EMBO J. 1996, 15, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Mukai, T.; Vargas-Rodriguez, O.; Englert, M.; Tripp, H.J.; Ivanova, N.N.; Rubin, E.M.; Kyrpides, N.C.; Söll, D. Transfer RNAs with novel cloverleaf structures. Nucleic Acids Res. 2017, 45, 2776–2785. [Google Scholar] [CrossRef] [PubMed]
- Mukai, T.; Englert, M.; Tripp, H.J.; Miller, C.; Ivanova, N.N.; Rubin, E.M.; Kyrpides, N.C.; Söll, D. Facile Recoding of Selenocysteine in Nature. Angew. Chem. Int. Ed. Engl. 2016, 55, 5337–5341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tharp, J.M.; Ehnbom, A.; Liu, W.R. tRNAPyl: Structure, function, and applications. RNA Biol. 2018, 15, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Polycarpo, C.; Ambrogelly, A.; Berube, A.; Winbush, S.M.; McCloskey, J.A.; Crain, P.F.; Wood, J.L.; Söll, D. An aminoacyl-tRNA synthetase that specifically activates pyrrolysine. Proc. Natl. Acad. Sci. USA 2004, 101, 12450–12454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doi, Y.; Ohtsuki, T.; Shimizu, Y.; Ueda, T.; Sisido, M. Elongation factor Tu mutants expand amino acid tolerance of protein biosynthesis system. J. Am. Chem. Soc. 2007, 129, 14458–14462. [Google Scholar] [CrossRef] [PubMed]
- Uhlenbeck, O.C.; Schrader, J.M. Evolutionary tuning impacts the design of bacterial tRNAs for the incorporation of unnatural amino acids by ribosomes. Curr. Opin. Chem. Biol. 2018, 46, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Xiong, H.; Reynolds, N.M.; Söll, D. Rationally evolving tRNAPyl for efficient incorporation of noncanonical amino acids. Nucleic Acids Res. 2015, 43, e156. [Google Scholar] [CrossRef] [PubMed]
- LaRiviere, F.J.; Wolfson, A.D.; Uhlenbeck, O.C. Uniform binding of aminoacyl-tRNAs to elongation factor Tu by thermodynamic compensation. Science 2001, 294, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Shepotinovskaya, I.; Uhlenbeck, O.C. tRNA residues evolved to promote translational accuracy. RNA 2013, 19, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Sun, S.B.; Furman, J.L.; Xiao, H.; Schultz, P.G. A versatile platform for single- and multiple-unnatural amino acid mutagenesis in Escherichia coli. Biochemistry 2013, 52, 1828–1837. [Google Scholar] [CrossRef] [PubMed]
- Schmied, W.H.; Elsässer, S.J.; Uttamapinant, C.; Chin, J.W. Efficient multisite unnatural amino acid incorporation in mammalian cells via optimized pyrrolysyl tRNA synthetase/tRNA expression and engineered eRF1. J. Am. Chem. Soc. 2014, 136, 15577–15583. [Google Scholar] [CrossRef] [PubMed]
- Serfling, R.; Lorenz, C.; Etzel, M.; Schicht, G.; Bottke, T.; Mörl, M.; Coin, I. Designer tRNAs for efficient incorporation of non-canonical amino acids by the pyrrolysine system in mammalian cells. Nucleic Acids Res. 2018, 46, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ambrogelly, A.; Gundllapalli, S.; Herring, S.; Polycarpo, C.; Frauer, C.; Söll, D. Pyrrolysine is not hardwired for cotranslational insertion at UAG codons. Proc. Natl. Acad. Sci. USA 2007, 104, 3141–3146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nozawa, K.; O’Donoghue, P.; Gundllapalli, S.; Araiso, Y.; Ishitani, R.; Umehara, T.; Söll, D.; Nureki, O. Pyrrolysyl-tRNA synthetase-tRNAPyl structure reveals the molecular basis of orthogonality. Nature 2009, 457, 1163–1167. [Google Scholar] [CrossRef] [PubMed]
- Fladischer, P.; Weingartner, A.; Blamauer, J.; Darnhofer, B.; Birner-Gruenberger, R.; Kardashliev, T.; Ruff, A.J.; Schwaneberg, U.; Wiltschi, B. A semi-rationally engineered bacterial pyrrolysyl-tRNA synthetase genetically encodes phenyl azide chemistry. Biotechnol. J. 2018, e1800125. [Google Scholar] [CrossRef] [PubMed]
- Meineke, B.; Heimgärtner, J.; Lafranchi, L.; Elsässer, S.J. Methanomethylophilus alvus Mx1201 provides basis for mutual orthogonal pyrrolysyl tRNA/aminoacyl-tRNA synthetase pairs in mammalian cells. ACS Chem. Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Willis, J.C.W.; Chin, J.W. Mutually orthogonal pyrrolysyl-tRNA synthetase/tRNA pairs. Nat. Chem. 2018, 10, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Borrel, G.; Gaci, N.; Peyret, P.; O’Toole, P.W.; Gribaldo, S.; Brugere, J.F. Unique characteristics of the pyrrolysine system in the 7th order of methanogens: Implications for the evolution of a genetic code expansion cassette. Archaea 2014, 2014, 374146. [Google Scholar] [CrossRef] [PubMed]
- Beranek, V.; Willis, J.C.W.; Chin, J.W. An evolved Methanomethylophilus alvus pyrrolysyl-tRNA synthetase/tRNA pair is highly active and orthogonal in mammalian cells. Biochemistry 2018. [Google Scholar] [CrossRef] [PubMed]
- Venkat, S.; Sturges, J.; Stahman, A.; Gregory, C.; Gan, Q.; Fan, C. Genetically incorporating two distinct post-translational modifications into one protein simultaneously. ACS Synth. Biol. 2018, 7, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Mukai, T.; Yamaguchi, A.; Ohtake, K.; Takahashi, M.; Hayashi, A.; Iraha, F.; Kira, S.; Yanagisawa, T.; Yokoyama, S.; Hoshi, H.; et al. Reassignment of a rare sense codon to a non-canonical amino acid in Escherichia coli. Nucleic Acids Res. 2015, 43, 8111–8122. [Google Scholar] [CrossRef] [PubMed]
- Krishnakumar, R.; Prat, L.; Aerni, H.R.; Ling, J.; Merryman, C.; Glass, J.I.; Rinehart, J.; Söll, D. Transfer RNA misidentification scrambles sense codon recoding. Chembiochem 2013, 14, 1967–1972. [Google Scholar] [CrossRef] [PubMed]
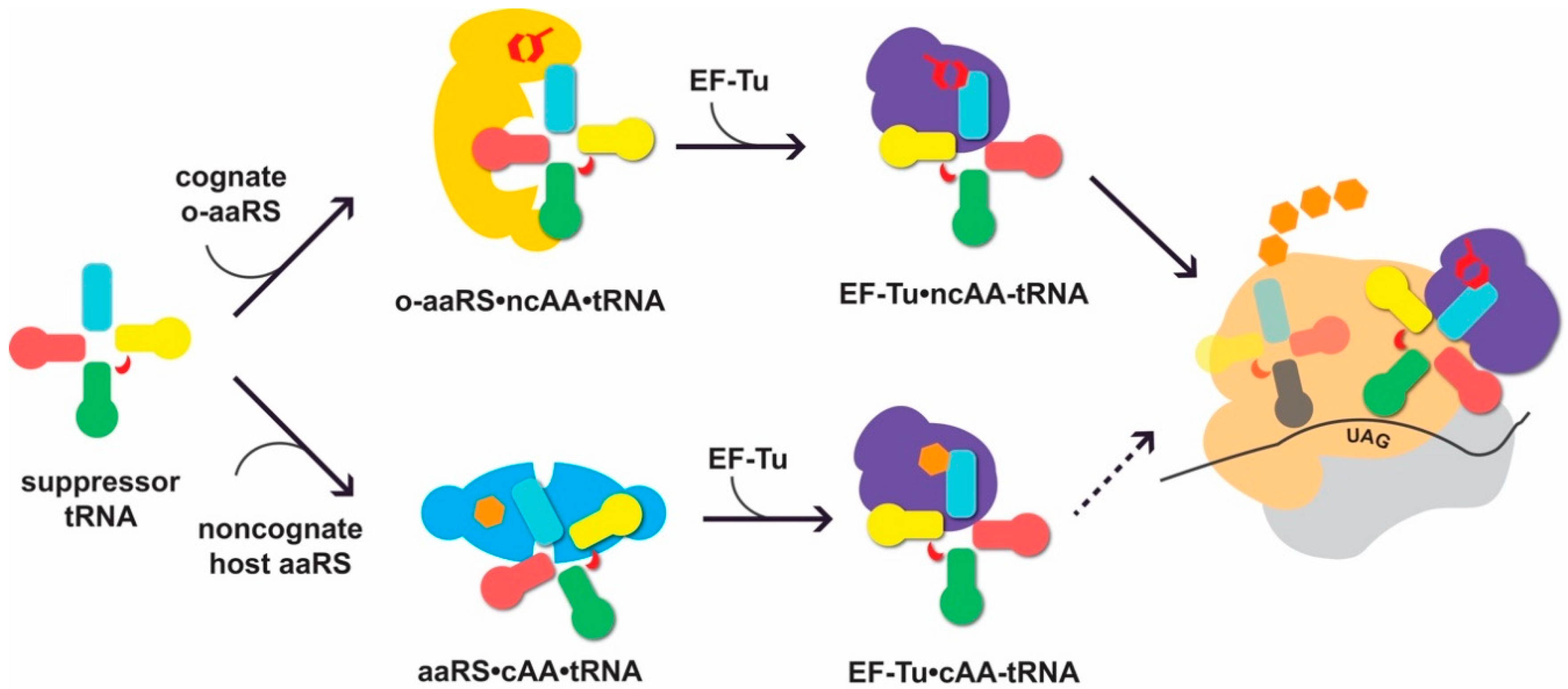
Figure 1.
Suppressor transfer RNAs (tRNAs) interact with cognate orthogonal aminoacyl-tRNA synthetases (o-aaRSs) and the translational machinery of the host. For successful non-canonical amino acid (ncAA) incorporation, the suppressor tRNA needs to be recognized by its cognate o-aaRS and charged with the cognate ncAA (up). When not orthogonal, the tRNA can be erroneously recognized by an endogenous noncognate aaRS and aminoacylated with a canonical AA (cAA; down). The formation of cAA-tRNA can lead to cAA incorporation at the ribosome in response to UAG (depicted as a dotted arrow). Elements of the tRNA secondary structure are shown in light blue (acceptor stem), pink (D-arm), green (anticodon arm), red (variable loop), and yellow (T-arm). The o-aaRS is shown in yellow, noncognate, endogenous aaRS in cyan, elongation factor EF-Tu in purple, and the large and small ribosomal subunit in tan and light grey, respectively. NcAA is depicted as a red hexagonal shape, while the natural AAs are given in orange. The position of the UAG codon is indicated.
Figure 1.
Suppressor transfer RNAs (tRNAs) interact with cognate orthogonal aminoacyl-tRNA synthetases (o-aaRSs) and the translational machinery of the host. For successful non-canonical amino acid (ncAA) incorporation, the suppressor tRNA needs to be recognized by its cognate o-aaRS and charged with the cognate ncAA (up). When not orthogonal, the tRNA can be erroneously recognized by an endogenous noncognate aaRS and aminoacylated with a canonical AA (cAA; down). The formation of cAA-tRNA can lead to cAA incorporation at the ribosome in response to UAG (depicted as a dotted arrow). Elements of the tRNA secondary structure are shown in light blue (acceptor stem), pink (D-arm), green (anticodon arm), red (variable loop), and yellow (T-arm). The o-aaRS is shown in yellow, noncognate, endogenous aaRS in cyan, elongation factor EF-Tu in purple, and the large and small ribosomal subunit in tan and light grey, respectively. NcAA is depicted as a red hexagonal shape, while the natural AAs are given in orange. The position of the UAG codon is indicated.

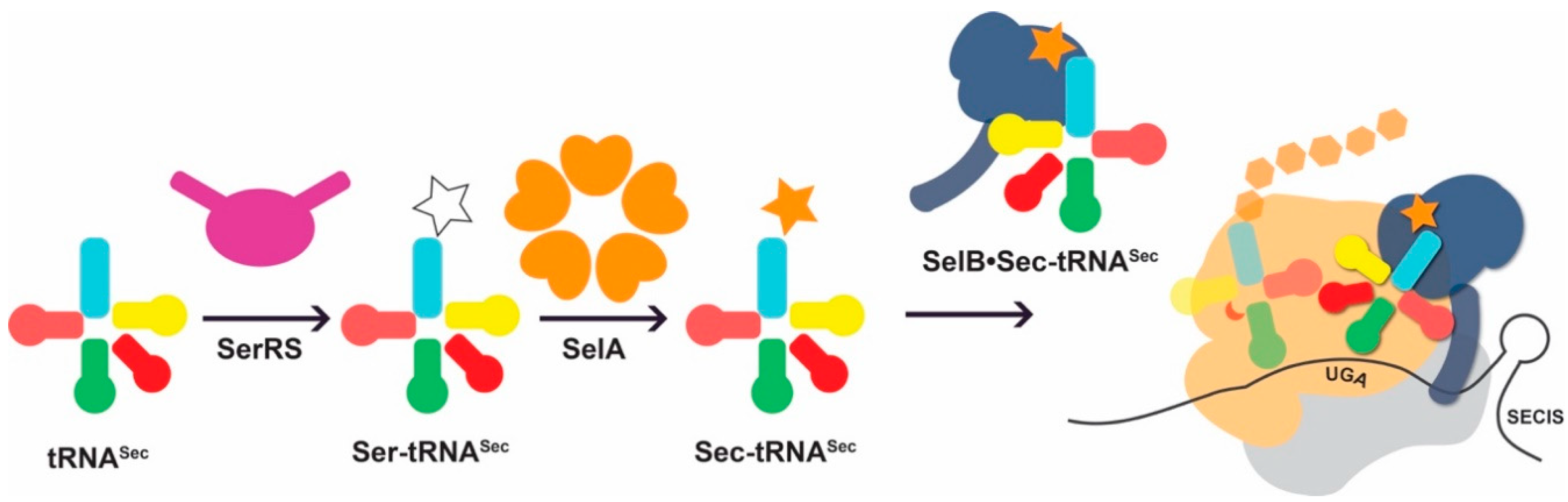
Figure 2.
Idiosyncratic features of the natural Sec-incorporation pathway. tRNASec is first misacylated with serine (white star) by seryl-tRNA synthetase (SerRS; purple). The intermediate, Ser-tRNASec is a substrate for selenocysteine synthase (SelA) which converts the Ser moiety to Sec (orange star). Sec-tRNASec is recognized by the Sec-specific elongation factor SelB (dark blue). In contrast to the general elongation factor EF-Tu, SelB approaches the ribosome bound to a Sec insertion sequence (SECIS), an RNA structure in its cognate mRNA. In this manner Sec-tRNASec is directed to bind an upstream UGA codon and deliver Sec to the growing polypeptide chain.
Figure 2.
Idiosyncratic features of the natural Sec-incorporation pathway. tRNASec is first misacylated with serine (white star) by seryl-tRNA synthetase (SerRS; purple). The intermediate, Ser-tRNASec is a substrate for selenocysteine synthase (SelA) which converts the Ser moiety to Sec (orange star). Sec-tRNASec is recognized by the Sec-specific elongation factor SelB (dark blue). In contrast to the general elongation factor EF-Tu, SelB approaches the ribosome bound to a Sec insertion sequence (SECIS), an RNA structure in its cognate mRNA. In this manner Sec-tRNASec is directed to bind an upstream UGA codon and deliver Sec to the growing polypeptide chain.

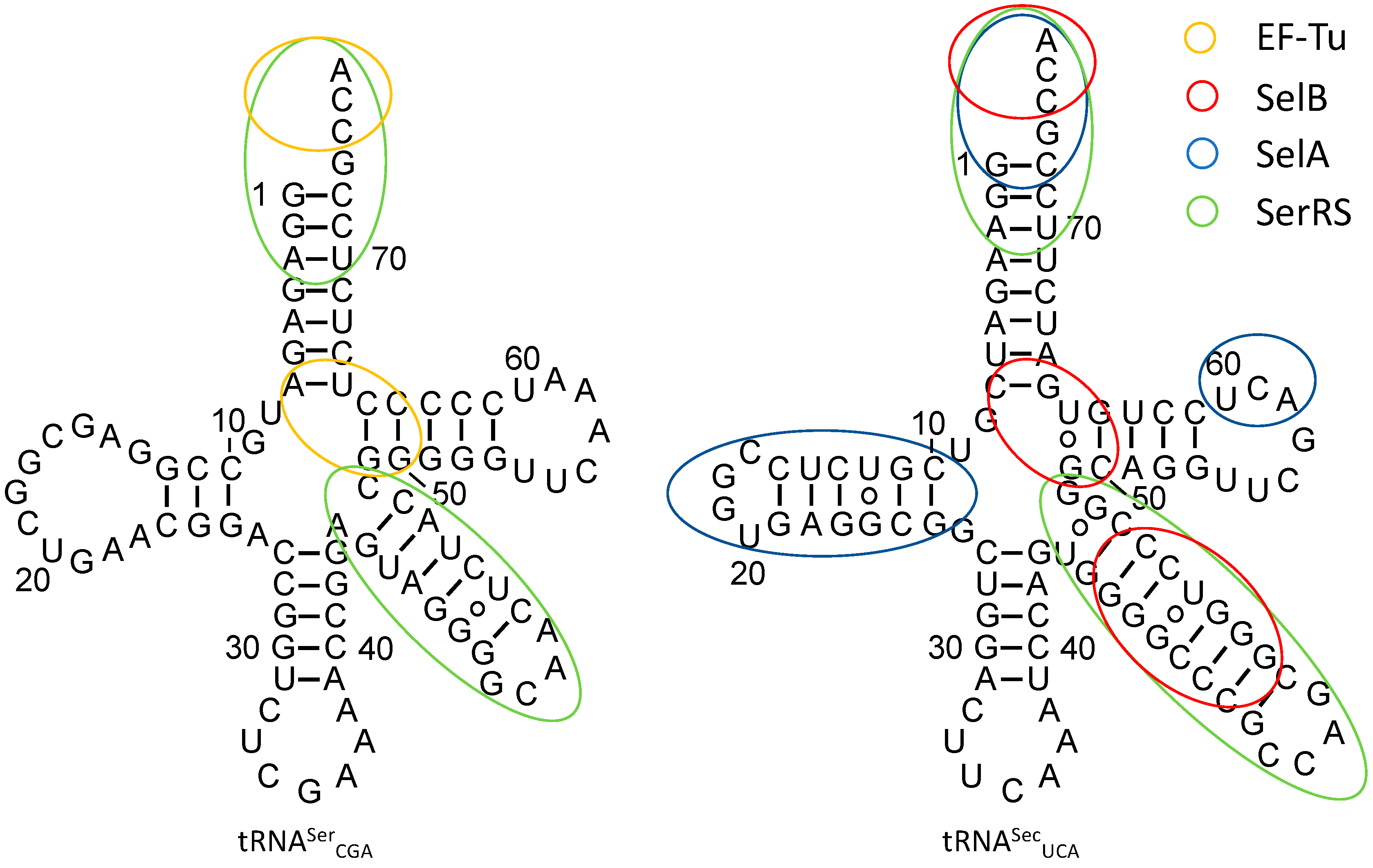
Figure 3.
Secondary structure of E. coli tRNASerCGA (left) and tRNASecUCA (right). Identity elements required for accurate recognition by EF-Tu, SelB, SelA, and SerRS are given in orange, red, blue, and green, respectively.
Figure 3.
Secondary structure of E. coli tRNASerCGA (left) and tRNASecUCA (right). Identity elements required for accurate recognition by EF-Tu, SelB, SelA, and SerRS are given in orange, red, blue, and green, respectively.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
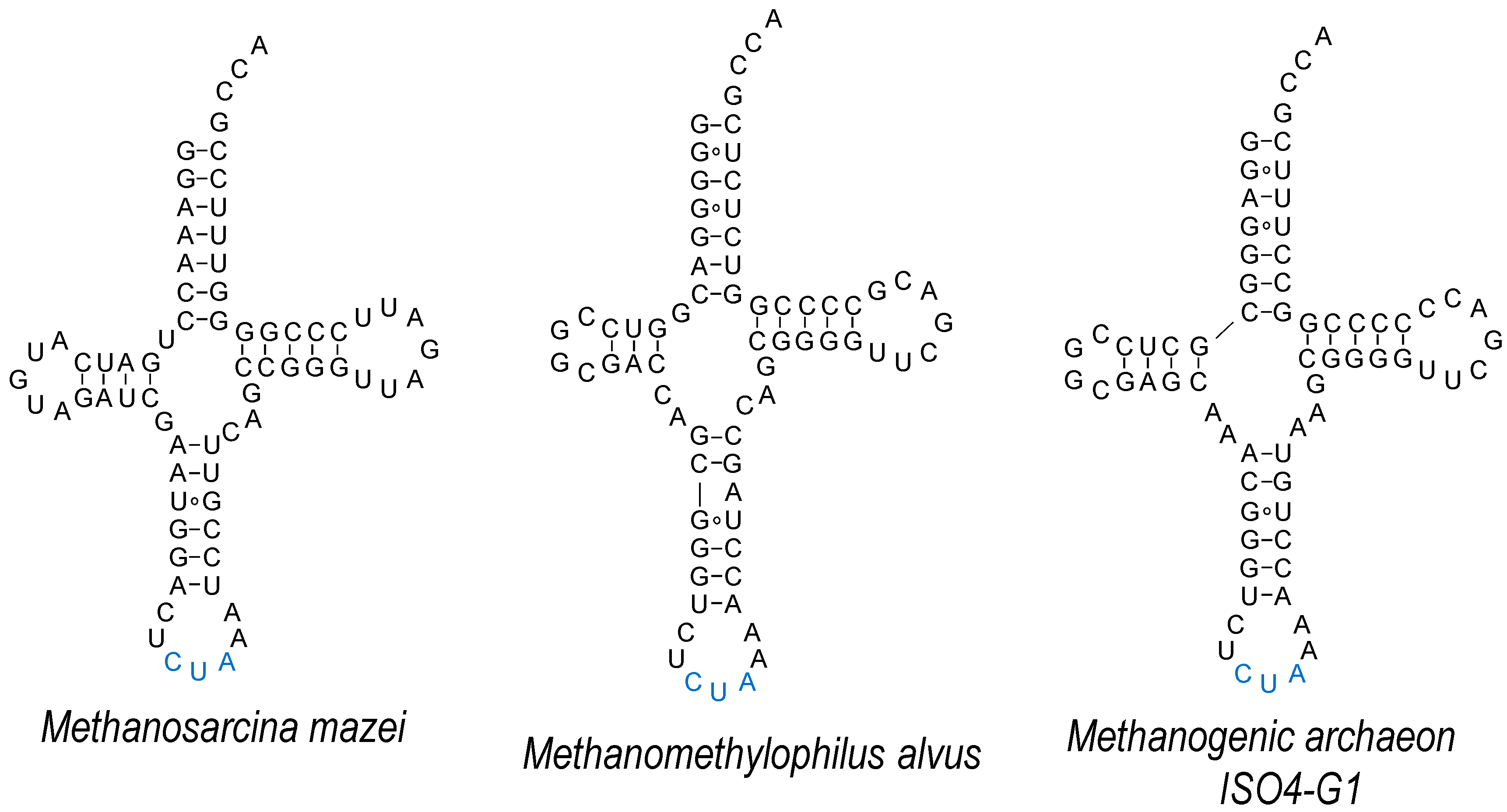{kind=link}
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hoffman, K.S.; Crnković, A.; Söll, D. Versatility of Synthetic tRNAs in Genetic Code Expansion. Genes 2018, 9, 537. https://doi.org/10.3390/genes9110537
AMA Style
Hoffman KS, Crnković A, Söll D. Versatility of Synthetic tRNAs in Genetic Code Expansion. Genes. 2018; 9(11):537. https://doi.org/10.3390/genes9110537
Chicago/Turabian StyleHoffman, Kyle S., Ana Crnković, and Dieter Söll. 2018. "Versatility of Synthetic tRNAs in Genetic Code Expansion" Genes 9, no. 11: 537. https://doi.org/10.3390/genes9110537
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.