Evaluating Short-Term Impacts of Forest Management and Microsite Conditions on Understory Vegetation in Temperate Fir-Beech Forests: Floristic, Ecological, and Trait-Based Perspective


Abstract
:1. Introduction
2. Materials and Methods
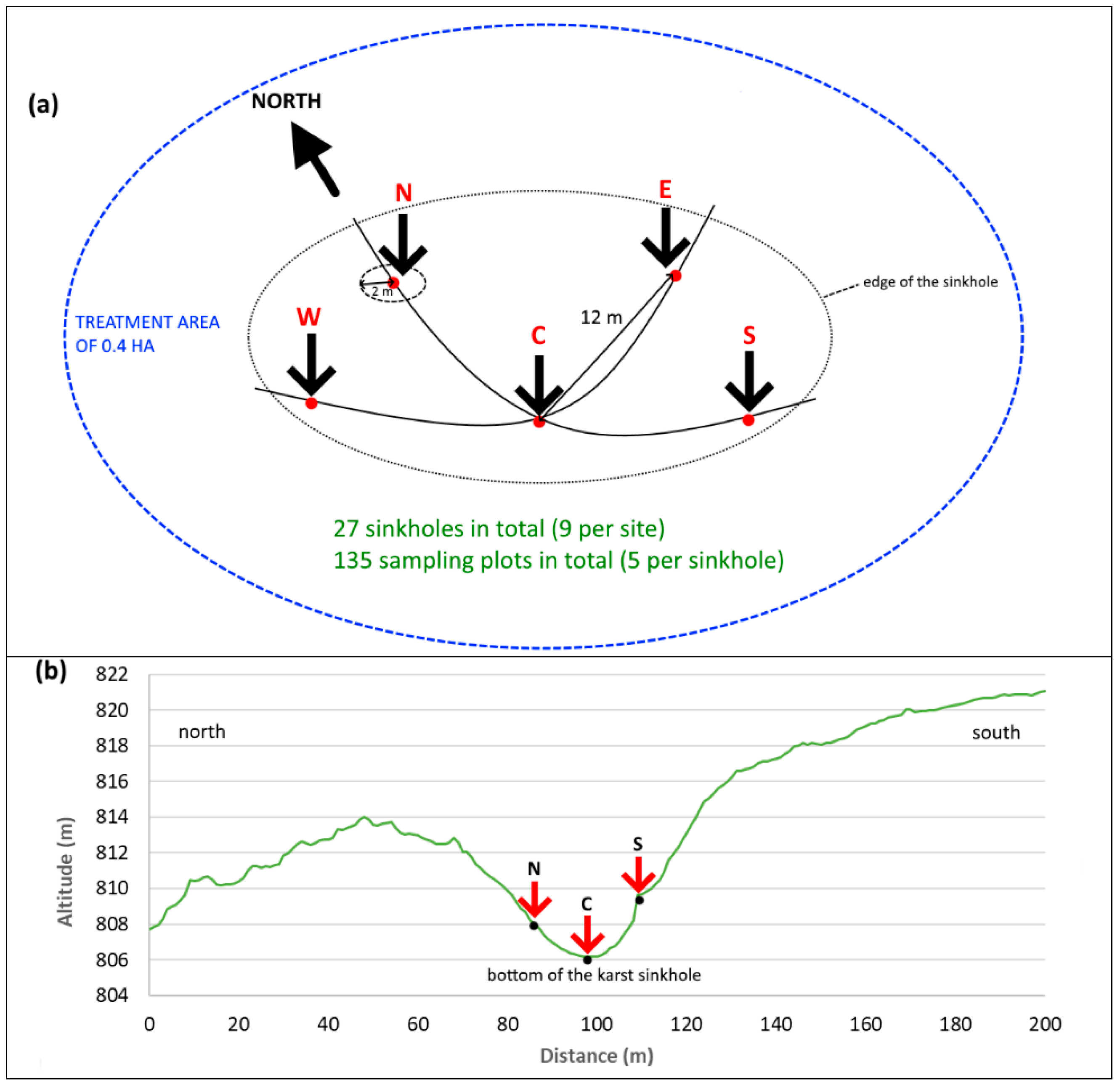
2.1. Study Design and Vegetation Sampling Methodology
2.2. Measurements of Post-Treatment Environmental Conditions
2.3. Data Processing and Analysis
3. Results
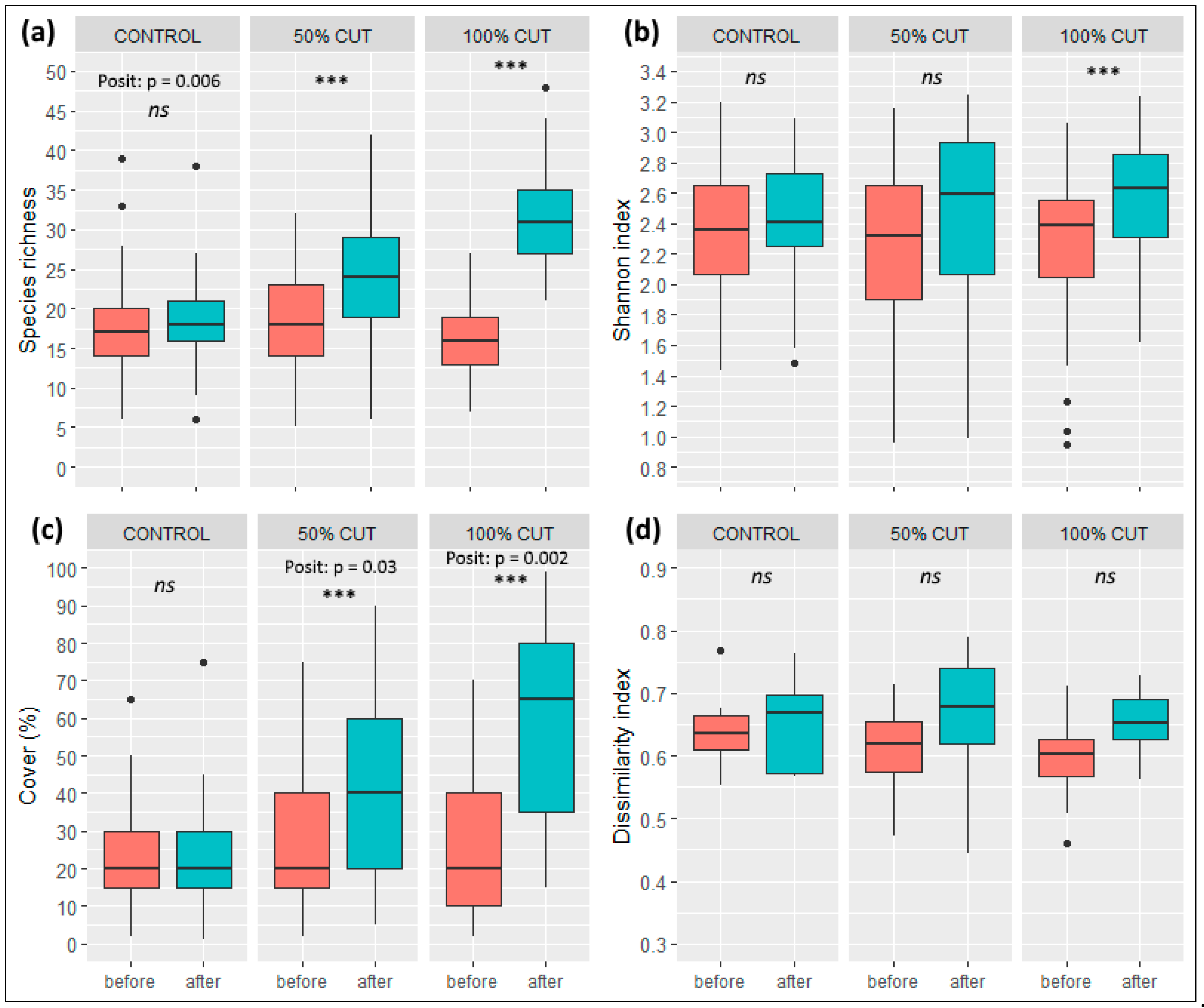
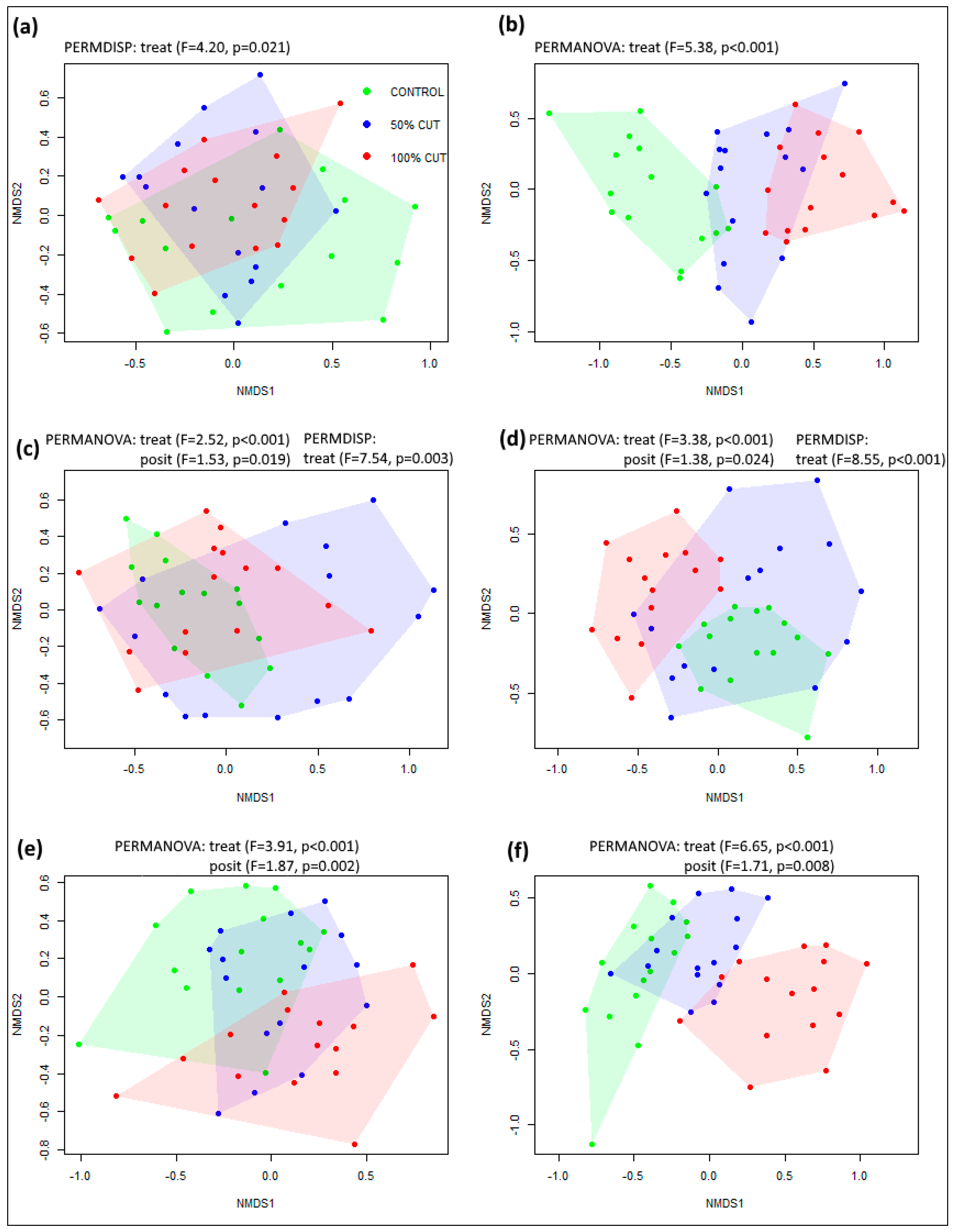
3.1. Changes in Species Diversity and Composition
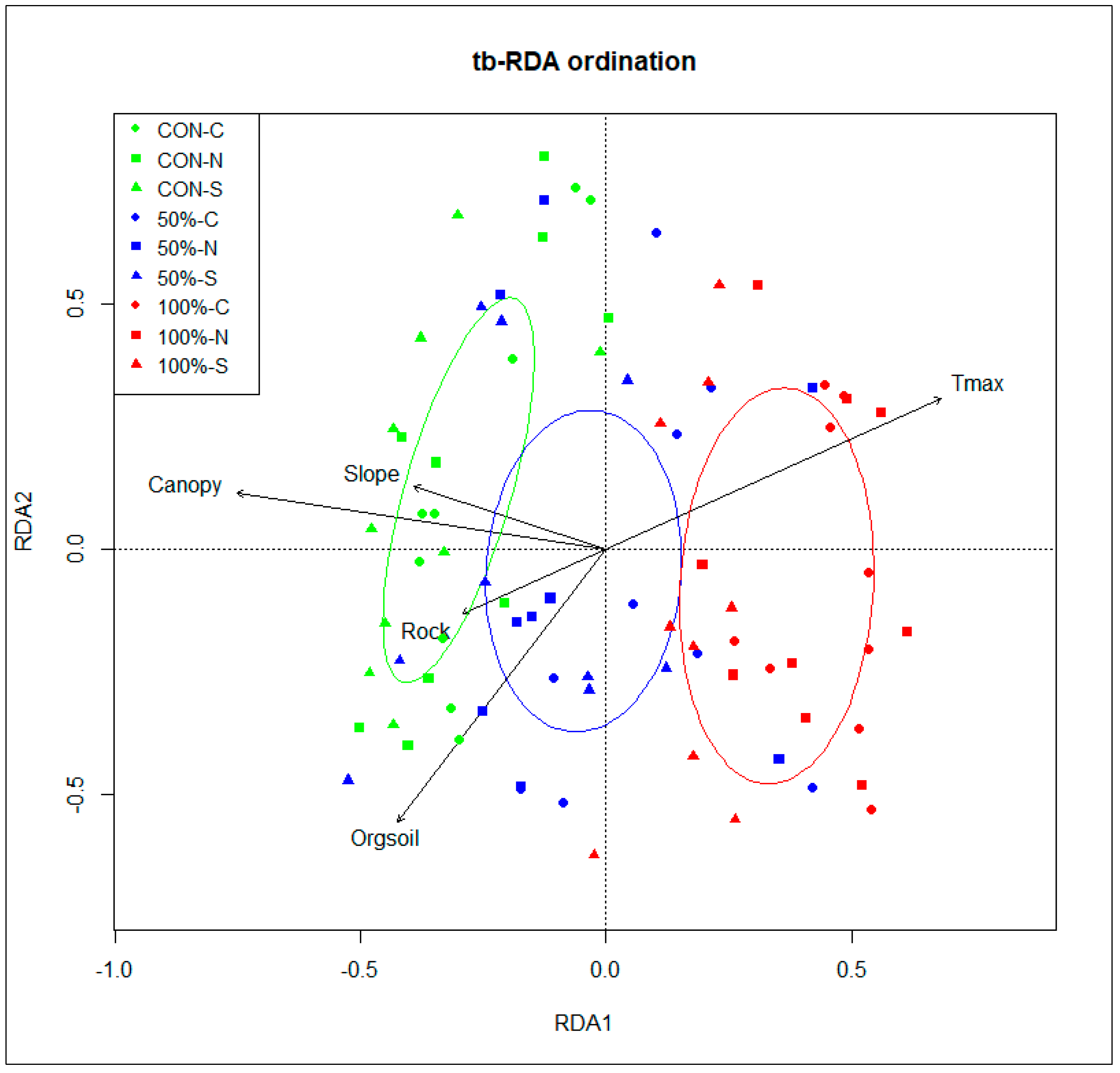
3.2. Linking Environmental Factors and Post-Disturbance Species Composition
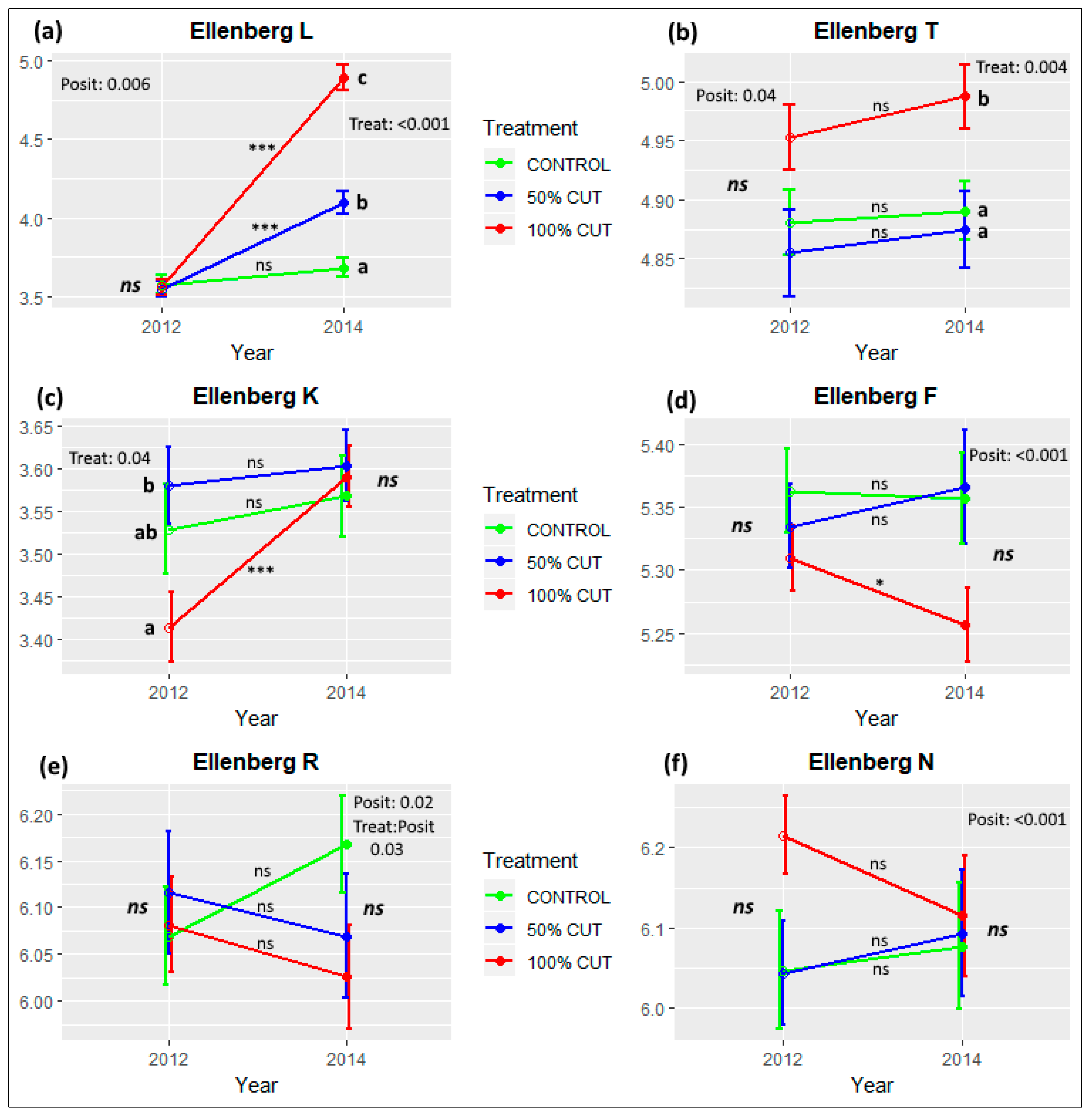
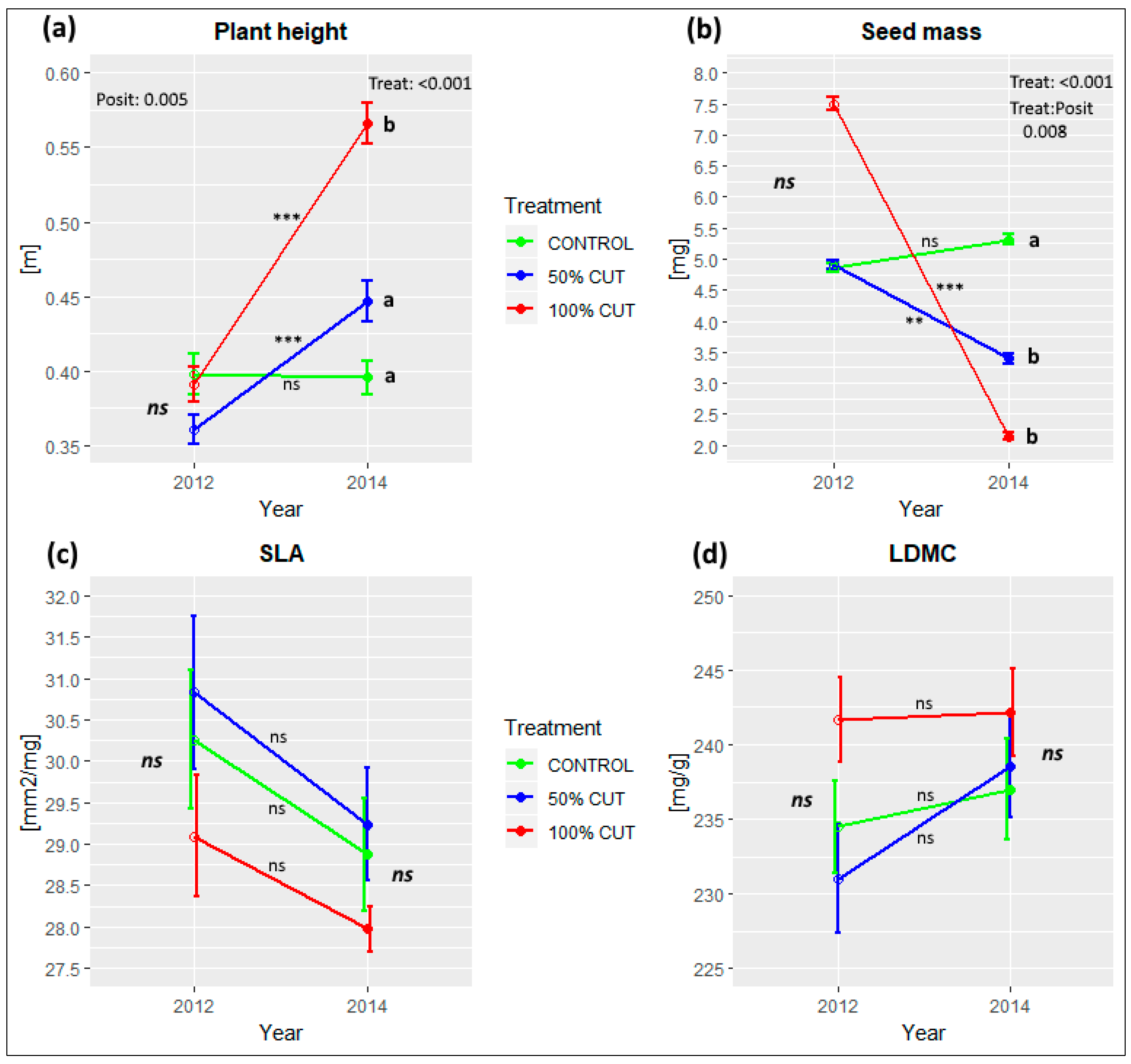
3.3. Effects of Treatment Intensity and Within-Sinkhole Position on EIVs and PFTs
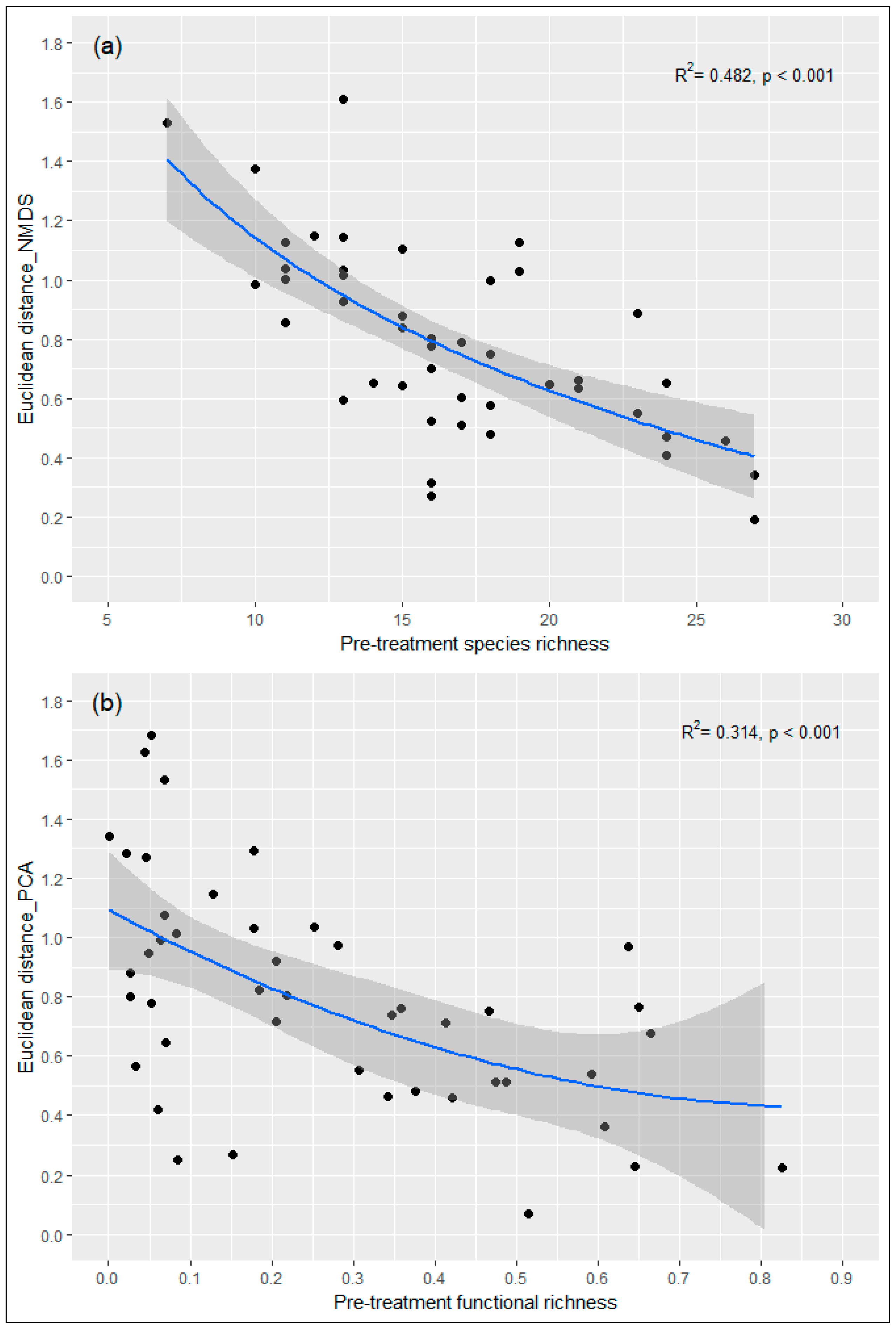
3.4. Influence of Pre-Treatment Richness on Post-Treatment Shifts
4. Discussion
4.1. Floristic Perspective
4.2. Ecological and Trait-Based Perspective
4.3. Initial Community Diversity as a Significant Determinant of Post-Disturbance Response
4.4. Potential Silvicultural Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lindenmayer, D.B.; Margules, C.R.; Botkin, D.B. Indicators of Biodiversity for Ecologically Sustainable Forest Management. Conserv. Biol. 2000, 14, 941–950. [Google Scholar] [CrossRef]
- Brunet, J.; Fritz, Ö.; Richnau, G. Biodiversity in European beech forests—A review with recommendations for sustainable forest management. Ecol. Bull. 2010, 53, 77–94. [Google Scholar]
- Lombardi, F.; Di Lella, S.; Altieri, V.; Di Benedetto, S.; Giancola, C.; Lasserre, B.; Kutnar, L.; Tognetti, R.; Marchetti, M. Early responses of biodiversity indicators to various thinning treatments in mountain beech forests. iForest 2018, 11, 609–618. [Google Scholar] [CrossRef]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Boch, S.; Prati, D.; Müller, J.; Socher, S.; Baumbach, H.; Buscot, F.; Gockel, S.; Hemp, A.; Hessenmöller, D.; Kalko, E.K.V.; et al. High plant species richness indicates management-related disturbances rather than the conservation status of forests. Basic Appl. Ecol. 2013, 14, 496–505. [Google Scholar] [CrossRef]
- Fedrowitz, K.; Koricheva, J.; Baker, S.C.; Lindenmayer, D.B.; Palik, B.; Rosenvald, R.; Beese, W.; Franklin, J.F.; Kouki, J.; Macdonald, E.; et al. Can retention forestry help conserve biodiversity? A meta-analysis. J. Appl. Ecol. 2014, 51, 1669–1679. [Google Scholar] [CrossRef]
- Dieler, J.; Uhl, E.; Biber, P.; Müller, J.; Rötzer, T.; Pretzsch, H. Effect of forest stand management on species composition, structural diversity, and productivity in the temperate zone of Europe. Eur. J. Forest Res. 2017, 136, 739–766. [Google Scholar] [CrossRef]
- Schmidt, W. Herb layer species as indicators of biodiversity of managed and unmanaged beech forests. For. Snow Landsc. Res. 2005, 79, 111–125. [Google Scholar]
- Paillet, Y.; Bergés, L.; Hjältén, J.; Ódor, P.; Avon, C.; Bernhardt-Römermann, M.; Bijlsma, R.-J.; De Bruyn, L.; Fuhr, M.; Grandin, U.; et al. Biodiversity Differences between Managed and Unmanaged Forests: Meta-Analysis of Species Richness in Europe. Conserv. Biol. 2010, 24, 101–112. [Google Scholar] [CrossRef]
- De Grandpré, L.; Boucher, D.; Bergeron, Y.; Gagnon, D. Effects of small canopy gaps on boreal mixedwood understory vegetation dynamics. Community Ecol. 2011, 12, 67–77. [Google Scholar] [CrossRef]
- Sabatini, F.M.; Burton, J.I.; Scheller, R.M.; Amatangelo, K.L.; Mladenoff, D.J. Functional diversity of ground-layer plant communities in old-growth and managed northern hardwood forests. Appl. Veg. Sci. 2014, 17, 398–407. [Google Scholar] [CrossRef]
- Horvat, V.; Biurrun, I.; García-Mijangos, I. Herb layer in silver fir-beech forests in the western Pyrenees: Does management affect species diversity? For. Ecol. Manag. 2017, 385, 87–96. [Google Scholar] [CrossRef]
- Aikens, M.; Ellum, D.; McKenna, J.J.; Kelty, M.J.; Ashton, M.S. The effects of disturbance intensity on temporal and spatial patterns of herb colonization in a southern New England mixed-oak forest. For. Ecol. Manag. 2007, 252, 144–158. [Google Scholar] [CrossRef]
- Burton, J.I.; Mladenoff, D.J.; Clayton, M.K.; Forrester, J.A. The roles of environmental filtering and colonization in the fine-scale spatial patterning of ground-layer plant communities in the north temperate deciduous forests. J. Ecol. 2011, 99, 764–776. [Google Scholar] [CrossRef]
- Bartels, S.F.; Chen, H.Y.H. Is understory plant species diversity driven by resource quantity or resource heterogeneity. Ecology 2010, 91, 1931–1938. [Google Scholar] [CrossRef]
- Royo, A.A.; Carson, W.P. On the formation of dense understory layers in forests worldwide: Consequences and implications for forest dynamics, biodiversity, and succession. Can. J. For. Res. 2006, 36, 1345–1362. [Google Scholar] [CrossRef]
- Kern, C.C.; Burton, J.I.; Raymond, P.; D’Amato, A.W.; Keeton, W.S.; Royo, A.A.; Walters, M.B.; Webster, C.R.; Willis, J.L. Challenges facing gap-based silviculture and possible solutions for mesic northern forests in North America. Forestry 2016, 90, 4–17. [Google Scholar] [CrossRef]
- Brosofske, K.D.; Chen, J.; Crow, T.R. Understory vegetation and site factors: Implications for a managed Wisconsin landscape. For. Ecol. Manag. 2001, 146, 75–87. [Google Scholar] [CrossRef]
- Durak, T. Changes in diversity of the mountain beech forest herb layer as a function of the forest management method. For. Ecol. Manag. 2012, 276, 154–164. [Google Scholar] [CrossRef]
- Márialigeti, S.; Tinya, F.; Bidló, A.; Ódor, P. Environmental drivers of the composition and diversity of the herb layer in mixed temperate forests in Hungary. Plant Ecol. 2016, 217, 549–563. [Google Scholar] [CrossRef] [Green Version]
- Ujházy, K.; Hederová, L.; Máliš, F.; Ujházyova, M.; Bosela, M.; Čiliak, M. Overstorey dynamics controls plant diversity in age-class temperate forests. For. Ecol. Manag. 2017, 391, 96–105. [Google Scholar] [CrossRef]
- Beatty, S.W. Influence of microtopography and canopy species on spatial patterns of forest understory plants. Ecology 1984, 65, 1406–1419. [Google Scholar] [CrossRef]
- Leuschner, C.; Lendzion, J. Air humidity, soil moisture and soil chemistry as determinants of the herb layer composition in European beech forests. J. Veg. Sci. 2009, 20, 288–298. [Google Scholar] [CrossRef]
- Duguid, M.C.; Ashton, M.S. A meta-analysis of the effect of forest management for timber on understory plant species diversity in temperate forests. For. Ecol. Manag. 2013, 303, 81–90. [Google Scholar] [CrossRef]
- Åström, M.; Dynesius, M.; Hylander, K.; Nilsson, C. Slope aspect modifies community responses to clear-cutting in boreal forests. Ecology 2007, 88, 749–758. [Google Scholar] [CrossRef]
- Diaci, J.; Adamic, T.; Rozman, A. Gap recruitment and partitioning in an old-growth beech forest of the Dinaric Mountains: Influences of light regime, herb competition and browsing. For. Ecol. Manag. 2012, 285, 20–28. [Google Scholar] [CrossRef]
- Vilhar, U.; Roženbergar, D.; Simončič, P.; Diaci, J. Variation in irradiance, soil features and regeneration patterns in experimental forest canopy gaps. Ann. For. Sci. 2015, 72, 253–266. [Google Scholar] [CrossRef]
- Diekmann, M. Species indicator values as an important tool in applied plant ecology—A review. Basic Appl. Ecol. 2003, 4, 493–506. [Google Scholar] [CrossRef]
- Dwyer, J.M.; Hobbs, R.J.; Mayfield, M.M. Specific leaf area responses to environmental gradients through space and time. Ecology 2014, 95, 399–410. [Google Scholar] [CrossRef]
- Amatangelo, K.L.; Johnson, S.E.; Rogers, D.A.; Waller, D.M. Trait-environment relationship remain strong despite 50 years of trait compositional change in temperate forests. Ecology 2014, 95, 1780–1791. [Google Scholar] [CrossRef]
- Burrascano, S.; Rosati, L.; Blasi, C. Plant species diversity in Mediterranean old-growth forests: A case study from central Italy. Plant Biosyst. 2009, 143, 190–200. [Google Scholar] [CrossRef]
- Bunn, W.A.; Jenkins, M.A.; Brown, C.B.; Sanders, N.J. Change within and among forest communities: The influence of historic disturbance, environmental gradients, and community attributes. Ecography 2010, 33, 425–434. [Google Scholar] [CrossRef]
- Belote, R.T.; Jones, R.H.; Wieboldt, T.F. Compositional stability and diversity of vascular plant communities following logging disturbance in Appalachian forests. Ecol. Appl. 2012, 22, 502–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, R.L.; Chen, H.Y.H.; Bartels, S.F.; Palik, B.J.; Prepas, E.E. Compositional stability of boreal understorey vegetation after overstorey harvesting across a riparian ecotone. J. Veg. Sci. 2015, 26, 733–741. [Google Scholar] [CrossRef]
- Kutnar, L.; Eler, K.; Marinšek, A. Effects of different silvicultural measures on plant diversity—The case of the Illyrian Fagus sylvatica habitat type (Natura 2000). iForest 2015, 9, 318–324. [Google Scholar] [CrossRef]
- Kermavnar, J.; Eler, K.; Marinšek, A.; Kutnar, L. Initial understory vegetation responses following different forest management intensities in Illyrian beech forests. Appl. Veg. Sci. 2019, 22, 48–60. [Google Scholar] [CrossRef]
- Eler, K.; Kermavnar, J.; Marinšek, A.; Kutnar, L. Short-term changes in plant functional traits and understory functional diversity after logging of different intensities: A temperate fir-beech forest experiment. Ann. For. Res. 2018, 61, 223–241. [Google Scholar] [CrossRef]
- Londo, G. The decimal scale for relevés of permanent quadrats. Vegetatio 1976, 33, 61–64. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package ‘vegan’—Community Ecology Package. 2019. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 5 May 2019).
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulißen, D. Zeigerwerte von Pflanzen in Mitteleuropa. 2. verbesserte und erweiterte Auflage. Scripta Geobotanica 1992, 18, 1–258. [Google Scholar]
- Pignatti, S.; Menegoni, P.; Pietrosanti, S. Valori di bioindicazione delle piante vascolari della Flora d’Italia. Bioindicator values of vascular plants of the Flora of Italy. Braun-Blanquetia 2005, 39, 3–95. [Google Scholar]
- Westoby, M. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant Soil 1998, 199, 213–227. [Google Scholar] [CrossRef]
- Thomson, F.J.; Moles, A.T.; Auld, T.D.; Kingsford, R.T. Seed dispersal distance is more strongly correlated with plant height than with seed mass. J. Ecol. 2011, 99, 1299–1307. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Diaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; ter Steege, H.; Morgan, H.D.; van der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- Herben, T.; Klimešová, J.; Chytrý, M. Effects of disturbance frequency and severity on plant traits: An assessment across a temperate flora. Funct. Ecol. 2018, 32, 799–808. [Google Scholar] [CrossRef]
- Kleyer, M.; Bekker, R.M.; Knevel, I.C.; Bakker, J.P.; Thompson, K.; Sonnenschein, M.; Poschlod, P.; van Groenendael, J.M.; Klimeš, L.; Klimešová, J.; et al. The LEDA traitbase: A database of life-history traits of the Northwest European flora. J. Ecol. 2008, 96, 1266–1274. [Google Scholar] [CrossRef]
- Lavorel, S.; Grigulis, K.; McIntyre, S.; Williams, N.S.G.; Garden, D.; Dorrough, J.; Berman, S.; Quétier, F.; Thébault, A.; Bonis, A. Assessing functional diversity in the field—Methodology matters! Funct. Ecol. 2008, 22, 134–147. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011; pp. 145–151. [Google Scholar]
- Andreson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar]
- Anderson, M.J.; Ellingsen, K.E.; McArdle, B.H. Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.W. Package ‘labdsv’—Ordination and Multivariate Analysis for Ecology. 2019. Available online: https://cran.r-project.org/web/packages/labdsv/labdsv.pdf (accessed on 21 June 2019).
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using Canoco 5; Cambridge University Press: Cambridge, UK, 2014; p. 373. [Google Scholar]
- Ter Braak, C.J.F. The analysis of vegetation-environment relationships by Canonical Correspondence Analysis. Vegetatio 1987, 69, 67–77. [Google Scholar] [CrossRef]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J. Constrained Ordination: Tutorial with R and vegan. 2012. Available online: http://cc.oulu.fi/~jarioksa/opetus/metodi/sessio2.pdf (accessed on 6 August 2019).
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effect Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009; 574p. [Google Scholar]
- Bartoń, K. “MuMIn”: Multi-Model Inference, R Package Version 1.43.6. 2019. Available online: https://cran.r-project.org/web/packages/MuMIn/MuMIn.pdf (accessed on 17 July 2019).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 21 April 2019).
- Kobal, M.; Bertoncelj, I.; Pirotti, F.; Dakskobler, I.; Kutnar, L. Using Lidar Data to Analyse Sinkhole Characteristics Relevant for Understory Vegetation under Forest Cover-Case Study of a High Karst Area in the Dinaric Mountains. PLoS ONE 2015, 10, e0122070. [Google Scholar] [CrossRef] [PubMed]
- Von Oheimb, G.; Härdtle, W. Selection harvest in temperate deciduous forests: Impact on herb layer richness and composition. Biodivers. Conserv. 2009, 18, 271–287. [Google Scholar] [CrossRef]
- Tinya, F.; Kovács, B.; Prättälä, A.; Farkas, P.; Aszalós, R.; Ódor, P. Initial understory response to experimental silvicultural treatments in a temperate oak-dominated forest. Eur. J. For. Res. 2019, 138, 65–77. [Google Scholar] [CrossRef]
- Aude, E.; Lawesson, J.E. Vegetation in Danish beech forests: The importance of soil, microclimate and management factors, evaluated by variation partitioning. Plant Ecol. 1998, 134, 53–65. [Google Scholar] [CrossRef]
- Weigel, R.; Gilles, J.; Klisz, M.; Manthey, M.; Kreyling, J. Forest understory vegetation is more related to soil than to climate towards the cold distribution margin of European beech. J. Veg. Sci. 2019, 30, 746–755. [Google Scholar] [CrossRef]
- De Groot, M.; Eler, K.; Flajšman, K.; Grebenc, T.; Marinšek, A.; Kutnar, L. Differential short-term response of functional groups to a change in forest management in a temperate forest. For. Ecol. Manag. 2016, 376, 256–264. [Google Scholar] [CrossRef]
- Michalová, Z.; Morrissey, R.C.; Wohlgemuth, T.; Bače, R.; Fleischer, P.; Svoboda, M. Salvage-Logging after Windstorm Leads to Structural and Functional Homogenization of Understory Layer and Delayed Spruce Tree Recovery in Tatra Mts., Slovakia. Forests 2017, 8, 88. [Google Scholar] [CrossRef]
- Gálhidy, L.; Mihók, B.; Hagyó, A.; Rajkai, K.; Standovár, T. Effects of gap size and associated changes in light and soil moisture on the understorey vegetation of a Hungarian beech forest. Plant Ecol. 2006, 183, 133–145. [Google Scholar] [CrossRef]
- Shipley, B.; Belluau, M.; Kühn, I.; Soudzilovskaia, N.A.; Bahn, M.; Penuelas, J.; Kattge, J.; Sack, L.; Cavender-Bares, J.; Ozinga, W.A.; et al. Predicting habitat affinities of plant species using commonly measured functional traits. J. Veg. Sci. 2017, 28, 1082–1095. [Google Scholar] [CrossRef]
- Reich, P.B.; Wright, I.J.; Cavender-Bares, J.; Craine, J.M.; Oleksyn, J.; Westoby, M.; Walters, M.B. The evolution of plant functional variation: Traits, spectra, and strategies. Int. J. Plant Sci. 2003, 164, 143–164. [Google Scholar] [CrossRef]
- Ampoorter, E.; Selvi, F.; Auge, H.; Baeten, L.; Berger, S.; Carrari, E.; Coppi, A.; Fotelli, M.; Radoglou, K.; Setiawan, N.N.; et al. Driving mechanisms of overstorey-understorey diversity relationships in European forests. Perspect. Plant Ecol. 2016, 19, 21–29. [Google Scholar]
- Haeussler, S.; Bedford, L.; Leduc, A.; Bergeron, Y.; Kranabetter, J.M. Silvicultural disturbance severity and plant communities of the southern Canadian boreal forest. Silva Fennica 2002, 36, 307–327. [Google Scholar] [CrossRef]
- Čater, M.; Diaci, J. Divergent response of European beech, silver fir and Norway spruce advance regeneration to increased light levels following natural disturbance. For. Ecol. Manag. 2017, 399, 206–212. [Google Scholar] [CrossRef]
- Čater, M.; Diaci, J.; Roženbergar, D. Gap size and position influence variable response of Fagus sylvatica L. and Abies alba Mill. For. Ecol. Manag. 2014, 325, 128–135. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dependent Variable | Model | Treatment | Position | Year | Treat: Posit | Treat: Year | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chi2 | p | R2LR | F | p | F | p | F | p | F | p | F | p | |
| Species diversity metrics | |||||||||||||
| Species richness | 184.96 | *** | 0.56 | 5.08 | * | 2.99 | * | 142.59 | *** | - | - | 46.18 | *** |
| Shannon index | 14.62 | *** | 0.16 | - | - | - | - | 14.95 | *** | - | - | - | - |
| Total cover | 145.88 | *** | 0.49 | 8.07 | ** | 6.27 | *** | 64.97 | *** | 2.17 | * | 28.82 | *** |
| Dissimilarity index | 4.43 | * | 0.08 | - | - | / | / | 4.46 | * | / | / | - | - |
| Tree sp. richness | 4.76 | * | 0.08 | - | - | - | - | 4.78 | * | - | - | - | - |
| Tree sp. cover | ns | ns | ns | - | - | - | - | - | - | - | - | - | - |
| Ellenberg indicator values (EIVs) | |||||||||||||
| Light (EIV-L) | 242.43 | *** | 0.62 | 21.52 | *** | 3.20 | * | 203.75 | *** | - | - | 59.69 | *** |
| Temperature (EIV-T) | 79.78 | *** | 0.41 | 6.28 | ** | 4.46 | ** | - | - | 2.79 | ** | - | - |
| Continentality (EIV-K) | 6.16 | * | 0.15 | - | - | - | - | 6.19 | * | - | - | - | - |
| Moisture (EIV-F) | 20.57 | *** | 0.54 | - | - | 5.27 | *** | - | - | - | - | - | - |
| Reaction (EIV-R) | 46.34 | *** | 0.49 | - | - | 4.91 | *** | - | - | 2.85 | ** | - | - |
| Nutrients (EIV-N) | 46.70 | *** | 0.46 | - | - | 7.62 | *** | - | - | - | - | - | - |
| Plant functional traits (CWMs) | |||||||||||||
| Plant height | 103.72 | *** | 0.44 | 5.85 | ** | 5.33 | *** | 53.02 | *** | - | - | 18.00 | *** |
| Seed mass | 155.15 | *** | 0.53 | - | - | - | - | 74.89 | *** | 2.91 | ** | 22.87 | *** |
| SLA | 20.28 | ** | 0.41 | - | - | 2.51 | * | 10.65 | ** | - | - | - | - |
| LDMC | ns | ns | ns | - | - | - | - | - | - | - | - | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kermavnar, J.; Marinšek, A.; Eler, K.; Kutnar, L. Evaluating Short-Term Impacts of Forest Management and Microsite Conditions on Understory Vegetation in Temperate Fir-Beech Forests: Floristic, Ecological, and Trait-Based Perspective. Forests 2019, 10, 909. https://doi.org/10.3390/f10100909
Kermavnar J, Marinšek A, Eler K, Kutnar L. Evaluating Short-Term Impacts of Forest Management and Microsite Conditions on Understory Vegetation in Temperate Fir-Beech Forests: Floristic, Ecological, and Trait-Based Perspective. Forests. 2019; 10(10):909. https://doi.org/10.3390/f10100909
Chicago/Turabian StyleKermavnar, Janez, Aleksander Marinšek, Klemen Eler, and Lado Kutnar. 2019. "Evaluating Short-Term Impacts of Forest Management and Microsite Conditions on Understory Vegetation in Temperate Fir-Beech Forests: Floristic, Ecological, and Trait-Based Perspective" Forests 10, no. 10: 909. https://doi.org/10.3390/f10100909